В царстве Chromista объединены грибы-водоросли и сходные с грибами нефотосинтезирующие гетеротрофы отдела Oomycota. Молекулярные исследования генов р-РНК малой и большой субъедниц рибосом показали, что грибы– водоросли (псевдогрибы) отдела Oomycota составляют единую линию эволюции с гетероконтными водорослями. Царство Хромиста включает бурые, золотистые, диатомовые гетероконтные водоросли. Еще в 1969 году Kreisel исключил эти грибы-водоросли из царства настоящих грибов. Для псевдогрибов характерны следующие особенности:
синтез лизина у всех идет одинаково, через аминоадипиновую кислоту (ДПА);
клеточные стенки содержат основные полисахариды: целлюлозу, глюканы, В 1-3, В 1-6, иногда – хитин;
у примитивных представителей в клетках нет настоящей стенки, но они покрыты слоями чешуек, продуцируемых аппаратом Гольджи. На поверхности клеток есть органеллы – ботросомы, связанные с эндомембранной системой и участвующей в транспорте веществ в сеть и обратно;
половой процесс – оогамный и (редко) изогамный;
бесполое размножение осуществляется зооспорами или апланоспорами. Зооспоры двужгутиковые, один из которых гладкий, а второй – перистый. Реже у псевдогрибов образуется один передний жгутик;
у некоторых наземных представителей роль зооспорангиев утрачена и они функционируют как конидии;
трофические связи: сапротрофы на органических остатках и паразиты водорослей, пресноводных и наземных растений;
псевдогрибы составляют единую линию эволюции с гетероконтными водорослями;
хромисты имеют монофилию.
Отдел Oomycota (Оомицеты)
Включает 700 видов с талломом в виде диплоидного одноклеточного мицелия, у немногих – ризомицелия. Отдел Oomycota выделен из царства Fungi (Eumycota) на основании отличия от настоящих грибов по морфологическим, биохимическим и ультраструктурным признакам. У всех представителей отдела половой процесс – своеобразная оогамия, при которой содержимое антеридия не дифференцируется на гаметы. Бесполое размножение осуществляется дву-жгутиковыми зооспорами с гетероморфными жгутиками – гладким и перистым, реже – одним передним жгутиком. Некоторые представители отдела образуют зооспорангии, функционирующие как конидии. Половой процесс завершается образованием телеоморфы – ооспоры.
Представители класса – облигатные внутриклеточные паразиты. Заболевания проявляются в виде опухолей на корнях растений. В цикле развития образуются покоящиеся, с толстой оболочкой споры, которые в оптимальных условиях образуют зооспоры с двумя разновеликими жгутиками. Класс состоит из одного порядка и одного семейства.
Порядок Plasmodiophorаlеs (Плазмодиофоровые)
Плазмодиофоровые включают в состав более 30 видов. Все они эндогенные паразиты, часто вызывающие гиперплазию и гипертрофию пораженных тканей растений. Плазмодии внутри гипертрофированных клеток хозяина сливаются и образуется многоядерный плазмодий. Затем плазмодий распадается на споры с тонкой оболочкой. После сгнивания пораженных растений споры освобождаются из клеток и прорастают в зооспорангий с одной зооспорой, имеющей два неравновеликих жгутика.
Семейство Plasmodiophorасеае (плазмодиофоровые)
В основу деления семейства на роды положены особенности строения покоящихся спор. Наиболее вредоносны два вида: Plasmodiophorа brassicae Wor. – возбудитель килы капусты и Spongospora subterranea Wallr. – возбудитель порошистой парши картофеля.
Рисунок 10 – Цикл развития возбудителя килы капусты P. brassicаe: 1 – гаплоидные зооспоры; 2 – диплоидная зооспора; 3 покоящиеся споры; 4 – гаметы; 5 – плазмодий.
При поражении P. brassicae на главном и боковых корнях растений капусты появляются наросты различной величины. Цвет нароста и корней вомногом совпадает. Внутри клеток опухоли плазмодии способны сливаться и образуют многоядерный плазмодий. Последний распадается на покоящиеся споры, которые после сгнивания опухолей прорастают в зооспорангий с одной двужгутиковой зооспорой. Характерной особенностью грибоподобной плазмодиофоры капустной является образование в клетках растения-хозяина не склеенных в кучки покоящихся спор.
На срезах опухолей больных корней видны плазмодии, распадающиеся на массу покоящихся спор, которые остаются в почве и могут сохранять жизнеспособность 5–6 лет. Весной они прорастают с образованием зооспор, которые в воде, по капилярам почвы передвигаются к корням капусты и проникают в клетки, образуя в них первичный плазмодий. Из плазмодиев вырастают зооспорангии с одножгутиковыми зооспорами. Зооспоры (+ и –) копулируют без слияния ядер и образовавшаяся двуядерная клетка проникает в корни растений, где образуется вторичный плазмодий с многочисленными ядрами. Плазмодий распадается на покоящиеся споры. При прорастании зооспор происходит увеличение клетoк растения в размере (гипертрофия) или массовое образование клеток в местах заражения (гиперплазия). В результате образуются объемистые опухоли (кила, рисунок 10) или бородавкообразные наросты – парша (рисунок 11).
Spongospora subterranea вызывает порошистую паршу картофеля только на клубнях, столонах, корнях. На них образуются бородавочки из ткани растения, которые посыхают и кожица разрывается в виде звездочки.
Царство Protozoa объединяет 4 отдела, среди которых лишь грибоподобные протисты (Plasmodiophoromycota) являются фитопатогенными. Они не имеют мицелия. Вегетативное тело их – плазмодий или псевдоплазмодий. Спороносные структуры – спорокарпы или сорокарпы, сходны со спороносными структурами грибов, что стало основанием для сближения протистов с грибами. Из псевдоплазмодия образуется сорокарп, клетки и выросты которого превращаются в споры. Споры бывают двужгутиковые и безжгутиковые. В клеточных стенках спор грибоподобных и в сорокарпах присутствует целлюлоза. В цикле развития грибоподобных имеются подвижные, с двумя гладкими жгутиками, и неподвижные формы. Это бесполое размножение у грибоподобных, а половой процесс – изогамия (с образованием диплоидного плазмодия). Кристы митохондрий в основном трубчатые, реже – дисковидные. Представители царства полифилетичны, но это требует дальнейших научных подтверждений (И. И. Сидорова, 2003). Наряду с фитопатогенными грибоподобными отдела Plasmodiophoromycota (плазмодиофоровые) в царство входят и наиболее распространенные представители отдела Myxomycota – настоящие слизевики.
Отдел Myxomycota (Слизевики)
Сюда отнесены типичные слизевики. Таллом слизевиков – диплоидный, многоядерный плазмодий (протопласт). В гаплоидно-диплоидном цикле развития мейоз* осуществляется перед формированием спорокарпа. Распространение происходит в основном гаплоидными спорами, образующимися в спорокарпе, клеточная стенка которых содержит целлюлозу. Половой процесс – изогамия – происходит при копуляции разнополых планогамет с образованием диплоидного амебоида. Пищеварение их эндогенное. В настоящее время отдел слизевиков как пограничный таксон также относится к животным. Отдел включает несколько классов, в том числе и самый многочисленный – Myxomycetes (Миксомицеты) с сапротрофным образом жизни представителей.
Класс Myxomycetes (Миксомицеты или Настоящие Слизевики)
В классе насчитывается более 1000 видов (Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л., 2006). Вегетативное тело их – многоядерный плазмодий (амебоид). Двигается плазмодий благодаря цитоплазматическим токам внутри и выростам в сторону движения.
При бесполом размножении образуются зооспоры с двумя неравновеликими гладкими жгутиками. Споры могут быть и без жгутиков. Половой процесс – копуляция планогамет. Представители класса обитают в почве, в гниющей древесине и плодовых телах шляпочных грибов и т. д.
Наиболее известен вид Physarum polycephalum с ярко-желтым или зеленовато – желтым плазмодием, развивающимся на гнилых плодовых телах (гименофорах) шляпочных грибов. На плазмодиях образуются спорангии с длинными извилистыми ножками. На спорангиях имеется перидий с известковыми узелками.
Отдел Plasmodiophoromycota (Плазмодиофоровые)
Отдел представлен грибоподобными организмами, вызывающими вредоносные заболевания растений и паразитирующие на гифах грибов-хозяев. Тело их – многоядерный диплоидный и гаплоидный плазмодий. Половой процесс – изогамия – слияние разнополых гаплоидных гамет с образованием диплоидного амебоида, который распадается на массу мелких, круглых покоящихся спор. Распространяются плазмодиофоровые покоящимися половыми спорами и бесполыми двужгутиковыми зооспорами с неравными гладкими жгутиками на переднем конце. Бесполое размножение некоторых представителей сопровождается образованием одножгутиковых зооспор.
В настоящее время общепризнана филогенетическая самостоятельность грибных организмов, отнесенных к Mycota. В природе существует около 1,5 млн видов грибов, из которых описано 75 тыс. (Хауксворт, 1992). Из них 5% являются патогенными для растений. Изучение грибов привело к необходимости разработки классификации их по группам со сходными эволюционными, морфологическими признаками.
По современным представлениям фитопатогенные грибы относятся к трем царствам живых организмов, из которых два царства представляют низшие грибы – грибоподобные протисты и грибы – водоросли, или псевдо-грибы, и одно – настоящие грибы. Систематика (номенклатура) грибов – распределение представителей царств на отделы, классы, подклассы, группы порядков, порядки, семейства, роды и виды. В основе построения систематики положен вид микроорганизма (species).
Современная систематика грибов основана на особенностях питания, морфологического сходства, эволюционных (филогенетических) связях между организмами. С появлением электронных микроскопов растровых (сканирующих) резко улучшилось рассматривание поверхностных компонентов на талломе и спороношениях грибов, а применение просвечивающей микроскопии позволило уточнить внутреннее строение объектов. В результате выявились детали клеток, недоступные при использовании световых микроскопов, что привело к расширению познания филогенетических построений биоты. Например, изучение ультраструктуры септы сложного строения у базидомицетов (долипоровой септы) позволило внести существенный вклад в выделение отдельным таксоном грибов класса устомицеты (Ustomycetes). В зооспорах оомицетов обнаружены вакуолярные структуры – К – тельца, которые отсутствуют у других грибов и грибоподобных организмов.
Химический состав и метаболизм имеют большое значение при выделении высоких таксонов. Так, клеточная оболочка оомицетов цементируется целлюлозой, а истинных грибов – хитином. Синтез аминокислот консервативен, однако у разных организмов имеются небольшие вариации, свойственные таксономическим связям. У псевдогрибов – оомицетов были обнаружены особенности синтеза лизина и скорости осаждения ферментов при биосинтезе аминокисот, что позволило отделить оомицеты от других классов и расположить в царстве грибов – водорослей, или псевдогрибов.
С 90-х годов прошлого века в систематике стали активно применяться молекулярные методы исследований грибов. Они основаны на анализе белков, РНК и ДНК. Разработанная на этой основе систематика получила название «Геносистематика» или «Молекулярная систематика». Современная систематика наиболее объективна потому, что позволяет определить нуклеотидные последовательности отдельных генов или участков ДНК, и судить о филогенетических связях таксонов любого уровня, их монофилии (таксоны, филогенетически происходящие от одной исходной предковой формы) и полифилии (таксоны, филогенетически происходящие от нескольких исходных предковых форм).
Применение молекулярных методов исследований грибов привело к коренным изменениям структуры их систематики. Со времен К. Линнея (в «домолекулярный» период) изучение грибов и их классификация базировались на познании морфологических, онтогенетических, физиолого-биохимических особенностей микроорганизмов. Однако такие научные подходы не позволяли выявить ключевые признаки для выделения основных эволюционных линий грибов: строение крист митохондрий, определение нуклеотидных последовательностей отдельных генов, или участков ДНК, накопление в эволюции нуклеотидов (принцип «молекулярных часов»), позволяющих подтвердить или опровергнуть монофилию тех или иных групп, на этой основе определить их эволюционные связи между собой и внести изменения в классификацию.
Методы геносистематики позволяют оценить возраст таксонов. Так, наиболее древней группой были хитридиомицеты. Через 100 млн лет возникли зигомицеты. Возникновение сумчатых и базидиальных грибов произошло позже – в девонский период (Белякова Г.А., Дьяков Ю.Т., Тарасов К.Л., 2006). Однако современные методы генной систематики имеют и недостатки. Так, молекулярные последовательности могут подвергаться эволюции, поэтому их закономерности, возможно, к настоящему времени не совпадают со структурой предковых форм.
Определение систематического положения возбудителя – обязательный этап в диагностике болезни, что позволяет биологически обосновать и разработать мероприятия по защите растений от болезней.
Таблица 1 — Схема классификации фитопатогенных грибоподобных организмов и настоящих грибов Mycota (Ainsworth and Bisby’s Dictionary of the fungi, 9th Edition, 2001)
Происхождение, особенности биологии грибов
По принятой в начале XXI века геносистематике классификация грибоподобных и настоящих грибов делится на 3 царства: Protozoa – слизевики или грибоподобные протисты; Chromista – хромисты, или грибы – водоросли, псевдогрибы; Fungi (Eumycota) – истинные (настоящие) грибы.
Грибы, предположительно, появились 900 млн лет назад независимо от растений и животных (Кутафьева Н. П., 2003). Грибы и грибоподобные организмы по различным гипотезам произошли от бурых, красных водорослей, амёб, жгутиконосов, цианобактерий. По строению, характеру обмена веществ и способу питания грибы занимают промежуточное положение между животными и растениями. Например, настоящие грибы – аскомицеты и базидиомицеты – с возможными предками (хитридиевыми грибами) происходили от жгутиконосных эукариот. Эукариоты потеряли жгутики в ходе приспособления к наземному образу жизни. Это позже произошло и с высшими растениями. Видимо, эволюция хитридиевых пошла по пути усовершенствования клетки, а настоящих грибов – по пути совершенствования морфологической структуры таллома.
Объединяет хитридиевые и настоящие грибы сходный синтез лизина – одной из аминокислот, которую они используют для синтеза белков цитоплазмы (Беккер З. Э., 1988). Этим данные грибы родственны между собой так же, как Protozoa (простейшие) и животные (Animale), зеленые водоросли (Chlorophyta) и растения (Plantae) [Э. Мюллер, В. Лёффлер, 1995; П. Рёйвн, Р. Эверт, С. Айкхорн, 1990].
Достаточно хорошо определились три линии эволюции у грибоподобных протистов и различных групп грибов. У них осуществлен переход к наземному образу жизни и изменена морфологическая структура таллома. Они имеют развитый осмогидротрофный способ питания и три его типа – сапротрофный, паразитарный и симбиотрофный. Третья линия – их репродукция с появлением огромного разнообразия видов и структур, позволяющих им осуществлять половое и бесполое размножение. Эти исследования привели в настоящее время к общему признанию положения о филогенетической самостоятельности грибных организмов. Уже описано достаточно много видов грибов из существующих на Земле (П. Рёйвн, Р. Эверт, С. Айкхорн, 1990).
Огромное биологическое разнообразие говорит о том, что эволюционно грибы – процветающие живые существа на планете. Изучение их невозможно без научно обоснованной классификации. На наш взгляд, наиболее обоснованной из ныне существующих классификаций (Э. Мюллер, В. Лёффер ,1995; П. Рёйвн, Р. Эверт, С. Айкхорн, 1990; М. В. Горленко, 1991 и др.) следует считать систематику фитопатогенных грибов по Ainsworth and Bisby’s гических справочников.
В существующих учебниках по общей фитопатологии систематика приведена по старому принципу, в соответствии со «Справочником по грибам», 7-е издание, 1985 г. За последнюю четверть века принципиальные изменения коснулись классов низших грибов – Chytridiomycetes, Oomycetes.
Специалистам в России 8-е и 9-е издания справочника малодоступны. Они есть только в личных библиотеках, в Интернете на сайте www.cabipublishing.org/bookshop/BookDisplay.asp. SubjectArea=& Subject=&PID=1529.
Предлагаемое учебное пособие, за некоторыми изменениями, соответствует «Методическим указаниям по систематике грибов и общей фитопатологии (для студентов факультета защиты и карантина растений)», изданным в 2004 г. Санкт-Петербургским государственным аграрным университетом. В них приведена систематика грибов в виде структуры крупных и мелких таксонов.
Cовременная систематика основана на особенностях :
морфологических;
онтогенетических;
ультраструктурных (строение крист митохондрий; наличие сформированных диктиосом, порового аппарата септ, зон контакта с растением – хозяином);
хемотаксономии (состава полисахаридов клеточных стенок, путей синтеза лизина и другие);
молекулярных (белков, РНК, ДНК);
определения нуклеотидных последовательностей отдельных генов или участков ДНК; определения накопленных в процессе эволюции замен нуклеотидов по принципу «молекулярных часов»;
изучения филогенетических связей таксонов с выявлением монофилии или полифилии.
Итак, в основу современной систематики положены: строение и состав клеточной стенки, особенности размножения грибов, физиологобиохимические свойства, ультраструктурные особенности клетки, филогенетическая и эволюционная связи между таксонами.
По-прежнему основным систематическим признаком и в современной классификации грибов является строение генеративных органов, образующих споры в результате полового процесса. В этой связи появились и некоторые названия таксонов высоких рангов. Например, грибы, продуцирующие зигоспоры, названы зигомицетами; образующие ооспоры – оомицетами, базидии с базидиоспорами – базидиомицетами, а сумки с сумкоспорами – сумчатыми.
Для грибоподобных протистов в основу систематики положены особенности строения и организации вегетативного тела, а также типов полового спороношения.
Согласно «Международному кодексу ботанической номенклатуры» основным таксоном является вид – species. В латинском названии отдела дописывается окончание – mycota, в классе – mycetes, подклассе – mycetidae, группе порядков – mycetes, порядке – ales, семействе – aceae.
После названия таксона, как правило, приводится фамилия автора, в скобках указывается автор, который ввёл для данного гриба видовой эпитет, а после скобок – автор, предложивший комбинацию родового и видового названия.
К фитопатогенным грибам относятся три группы организмов: грибоподобные протисты (царство Protozoa, Protista), псевдогрибы или грибы – водоросли (царство Chromysta) – хромисты и царство настоящих (истинных грибов) – Fungi (Eumycota), (таблица 2).
Таблица 2 – Kлассификация фитопатогенных грибоподобных организмов царств Protozoa, Chromista и низших грибов царства Eumycota, (Dictionary of the fungi, 9th Edition, 2001)
У грибоподобных и настоящих грибов различают два типа размножения: вегетативное и репродуктивное. Репродуктивное размножение происходит бесполым и половым путем. Вегетативное размножение грибов осуществляется частями гиф мицелия, или его видоизменениями – оидиями, бластоспорами, хламидоспорами, геммами и другими (рисунок 4). Все они, попав в благоприятные условия, могут дать начало новому мицелию.
Оидии (греч. oon – яйцо, овальная клетка), или артроспоры, образуются в результате фрагментации конидиеносца, гифы или всего мицелия на отдельные эллипсоидальные или шаровидные клетки с тонкой оболочкой. Они недолговечны и способствуют распространению инфекции в период вегетации. Оидии образуют грибы из родов Endomyces, Oidium, Oidiodendron и другие. Бластоспоры (греч. blastos – росток + spora – семя) образуются в результате почкования гиф (почкующийся мицелий у дрожжевых грибов рода Saccharomyces) или спор (почкующиеся споры у голосумчатых грибов рода Taphrina). При этом на клетке мицелия или споре образуется маленький вырост, который постепенно увеличивается в размере, а после окончания роста отделяется от материнской клетки и снова начинает почковаться. В результате почкования формируется цепочка бластоспор, которые называются псевдомицелием. Разнообразные формы вегетативного размножения способствуют увеличению численности и длительному сохранению грибов при неблагоприятных условиях внешней среды.
Оно происходит посредством особых спор, имеющих специфические названия зооспор, спорангиоспор и конидий. По отношению к образующим их органам бывают эндогенными – зооспоры, спорангиоспоры (грибоподобные), пикноспоры (настоящие грибы) и экзогенными – конидии (у грибоподобных и настоящих грибов). Зооспоры (греч. spora – семя + zoon – животное) – одноклеточные, одноядерные подвижные споры с одним или двумя жгутиками, сохраняющими жизнеспособность только в воде (рисунок 5). Образуются в специальных шаровидных или грушевидных вместилищах – зооспорангиях (греч. zoon – животное + spora – семя + angeion – сосуд), которые формируются на концах особых гиф – зооспорангиеносцах. При созревании оболочка зооспорангия разрывается, зооспоры освобождаются и при помощи жгутиков активно передвигаются в воде. Это простейшая форма бесполого размножения грибоподобных (рода Plasmo-diophora – класс Plasmodiophoromycetes; рода Synchytrium, Olpidium – класс Chytridiomycetes; родов Saprolegnia, Plasmopara, Phytophthora – класс Oomycetes). Спорангиоспоры – одноклеточные неподвижные споры, образующиеся в шаровидных вместилищах – спорангиях (греч. spora – семя + angeion – сосуд) на особых гифах мицелия – спорангиеносцах. Спорангиоспоры имеют твердую оболочку. Освобождаясь через разрывы спорангия, они распространяются с потоками воздуха на большие расстояния. Данный тип бесполого размножения характерен для грибов порядка Mucorales (класс Zygomycetes).
Рисунок 5 – Органы бесполого репродуктивного размножения грибов: 1 – зооспорангии (а) с зооспорами (б) у грибов рода Olpidium; 2–типы строения жгутиков: (а) – задний бичевидный, (г) – передний перистый, и зооспора с двумя типами жгутиков – перистым и бичевидным; 3 –спорангиеносец (а), спорангии со спорангиоспорами (в) у грибов рода Mucor; 4 – конидиеносец (а) и конидии (б) у грибов рода Verticillium.
Эндогенные споры, характерны для грибоподобных хромистов и протистов, четко демонстрируют их связь с водным образом жизни. Переход к жизни на суше привел к эволюции зооспорангия в спорангий, подвижных зооспор – в неподвижные спорангиоспоры. Дальнейшее совершенствование грибов при жизни на суше привело к возникновению экзогенного спорообразования – конидиеносцев с конидиями. Конидии (греч. konia – пыль + eidos – вид) – споры, образующиеся на конидиеносцах. Конидиеносцы – это простые или более или менее дифференцированные ответвления гиф, на которых возникают клетки, образующие конидии. Конидии и конидиеносцы очень разнообразны по размерам, форме, окраске и строению, а также по характеру развития и размещения. Конидии бывают одно- и многоклеточные, шаровидные, округлые, булавовидные, нитевидные и другие, а так же бесцветные и окрашенные. Конидиеносцы бывают простыми, напоминающими гифы, или сложными, с различным по сложности ветвлением. Конидиеносцы могут располагаться на мицелии одиночно или формировать агрегации в виде коремий, пионнот, спородохий, а также образовывать тесный слой на ложе и в пикниде (рисунок 6).
Рисунок 6 – Способы формирования конидиального спороношения: 1 – одиночные конидиеносцы (а – род Ramularia, б – род Cephalosporium, в – род Arthrobotrys, г – род Embellisia, д – род Alternaria); 2 – спородохии (род Tubercularia); 3 – коремия; 4 – ложе (род Marssonina); 5 – пикнида (род Diplodia).
Коремии – это пучки плотно сближенных, обычно склеенных, а иногда и сросшихся друг с другом конидиеносцев. В результате этого образуется компактная колонка, на вершине которой на веточках конидиеносцев синхронно развиваются конидии. Коремии характерны для представителей порядка Hyphomycetales (семейство Tuberculariaceae, класс Hyphomycetes). Спородохии – это конидиеносцы с конидиями, собранные в подушечки на поверхности выпуклого сплетения гиф. Если конидиальное спороношение такого типа имеет слизистую или желеобразную консистенцию, а в основании более рыхлое сплетение гиф мицелия, их называют пионнотами. Такой тип спороношения характерен для представителей порядка Hyphomycetales. Ложе – тесный слой конидиеносцев с конидиями на более или менее плоском сплетении гиф. Ложе может располагаться на поверхности или внутри питающего растения. Оно часто покрыто покровными тканями (кутикулой или эпидермисом) и раскрывается к моменту созревания конидий. Ложе характерно для представителей порядка Melanconiales (класс Coelomycetes). Пикниды – шарообразные или кувшиновидные вместилища, с плотной, светлой или темной оболочкой, состоящей из сплетения мицелия, с узким отверстием наверху – устьицем. Внутри пикниды (эндогенно) образуются плотным слоем конидиеносцы, на которых формируются конидии, выходящие затем из пикниды через устьице или трещины в оболочке, часто в массе слизи. Пикниды характерны для представителей порядка Sphaeropsidales (класс Coelomycetes). У грибов можно проследить постепенный переход от эндогенного бесполого размножения с помощью спорангиoспор к экзогенному бесполому размножению с помощью конидий. Форма бесполого размножения характерна преимущественно для настоящих грибов, но встречается и у низших с более высокой организацией. У многих фитопатогенных грибов в период вегетации возникает несколько генераций бесполого репродуктивного спороношения. Эти споры способствуют массовому распространению природного инокулюма и повторному заражению растений.
Половое размножение
Половое размножение грибов заключается в слиянии мужских и женских половых клеток – гамет (греч. gametes – супруг, gamete – супруга), в результате чего образуется зигота (греч. zygote – соединение в пару). При образовании зиготы ядра гаплоидных гамет сливаются, число хромосом удваивается, т. е. наступает диплоидная фаза. В дальнейшем, после репродуктивного деления диплоидного ядра, наступает гаплоидное состояние. В половом процессе различают три основные фазы: – плазмогамия – слияние отцовской и материнской клетки; – кариогамия – слияние ядер; – редукция – деление ядер. Эти фазы протекают иногда очень быстро одна за другой, а иногда – растягивающиеся на период, охватывающий почти всю жизнь данного организма. У низших псевдогрибов и низших грибов – зигомицетов после оплодотворения клетка переходит в покоящееся состояние (ооспору, зигоспору). У настоящих грибов после оплодотворения клетка не переходит в состояние покоящейся споры, а развивается в дикариотичный, или диплоидный мицелий. Впоследствии на этом мицелии формируются аски (сумки) с аскоспорами (сумкоспорами) – у аскомицетов, и базидии с базидиоспорами – у базидиомицетов. Известны следующие типы полового размножения грибов: – планогамия, оогамия и зигогамия – у низших организмов (протистов, хромистов) и настоящих грибов (хитридиомицетов, зигомицетов); – гаметангиогамия и соматогамия – у настоящих грибов. Планогамия – слияние подвижных и разнополых гамет, ослабленных условиями окружающей среды или голоданием. В результате копуляции образуется дикарион – подвижная планозигота, снабженная жгутиком. Она внедряется в ткани растения, теряет жгутик, становится неподвижной, покрывается плотной оболочкой и превращается в покоящуюся спору – цисту. Циста легко переносит неблагоприятные условия внешней среды и после периода покоя прорастает как зооспорангий. Половой процесс типа планогамии наблюдается у представителей классов Plasmodiophoromycetes и Chytridiomycetes. Планогамия бывает: изогамная (греч. isos – одинаковый + gamos – брак) – копуляция двух подвижных, одинаковых по форме и размеру гамет, но разных в половом отношении; гетерогамная (греч. heteros – другой + gamos – брак) – слияние подвижных разнополых гамет неодинаковой величины; оогамная (греч. ооn – яйцо + gamos – брак) – слияние мужской одножгутиковой подвижной гаметы с женской одноядерной шаровидной гаметой-яйцеклеткой, свободно лежащей в оогонии (рисунок 7). Оогамия – слияние неподвижных, различных по форме и строению половых клеток: женской – оогония (греч. ооn – яйцо + gonos – рождение) с мужской – антеридием (греч. аntheros – цветущий). В оогонии может быть одна яйцеклетка (порядок Peronosporales) или несколько (порядок Saprolegniales). После слияния содержимого половых клеток образуется покоящаяся спора – ооспора, окруженная плотной многослойной оболочкой. После периода покоя ооспора прорастает в зародышевый зооспорангий (Albugo), или в короткую гифу с зародышевым зооспорангием на конце (Plasmopara), или в мицелий (Peronospora). Оогамия характерна для представителей класса Oomycetes.
Рисунок 7 – Половое репродуктивное размножение у некоторых настоящих грибов и псевдогрибов: 1 – планогамия (изогамная (а), гетерогамная (б), оогамная (в) у хитридиомицетов; 2 – зигогамия: последовательные стадии формирования зигоспоры (а) и ее прорастание (б); 3 – оогамия, оплодотворение оогония (а) с помощью антеридия (б) у оомицетов; ооспора (в).
Зигогамия – слияние двух внешне одинаковых многоядерных клеток разнополых мицелиев. При этом два гаметангия растут навстречу друг другу, в месте соприкосновения они отделяют по одной многоядерной клетке, оболочка между ними в результате этого растворяется и содержимое их сливается, а ядра образуют множество дикарионов. Образовавшаяся клетка – зигоспора, покрывается снаружи многослойной, пигментированной, бугорчатой оболочкой. Прорастает зигоспора в мицелий с первичным (зародышевым) спорангием. Зигогамия характерна для мукоровых грибов (класс Zygomycetes). Гаметангиогамия (греч. gametes – супруг + angeion – сосуд + gamos – брак) – оплодотворение женской половой клетки – аскогона мужской клеткой – антеридием с помощью трихогины (рисунок 8). Многоядерный антеридий формируется на мужской гифе, на женской – многоядерный аскогон с узкой вытянутой клеткой на вершине – трихогиной. При оплодотворении в оболочке трихогины образуется отверстие, через которое содержимое антеридия переливается в аскогон. В результате гаметангиогамии образуются сумки, или аски (лат. asc – сумка) с четным количеством, чаще с восемью, аскоспорами. Гаметангиогамия протекает у большинства представителей отдела Ascomycоtа. Формирование аскоспор происходит следующим образом. После плазмогамии из аскогона отрастают аскогенные гифы, которые разделяются на клетки с двумя ядрами в каждой. В каждой клетке вначале проходит простое деление ядер. Затем они сливаются, образуя диплоидное ядро, в котором происходит редукционное и два простых деления. Из верхней части крючковидного выроста формируется аск (сумка) с восемью аскоспорами. Грибы класса Hemiascomycetes (голосумчатые) не образуют плодовых тел. Сумки формируются на мицелии из зиготы, без образования аскогенных гиф. У представителей класса Euascomycetes (плодосумчатые) сумки заключены в плодовые тела: клейстотеции, перитеции и апотеции. Группа порядков Loculo-ascomycetes характеризуется отсутствием настоящих плодовых тел. Аски формируются в особых полостях (локулах) мицелиальных стром – аскостромах или псевдотециях (рисунок 9). Соматогамия (греч. soma – тело + gаmos – брак) слияние двух клеток вегетативного гаплоидного мицелия, выросшего из базидиоспор (отдел Basidio – mycota) (см. рисунок 8). У базидиальных грибов в цикле развития преобладает дикариотическое и диплоидное состояние. Переход от гаплоидного состояния к дикариотическому может происходить в пределах одного мицелия. Такое явление называется гомоталлизмом (греч. homos – одинаковый + thallus – тело).
Рисунок 8 – Половое репродуктивное размножение настоящих грибов: 1 – половой процесс и развитие сумок у аскомицетов (антеридий (а) и аскогон (б) с трихогиной; развитие сумок (в); 2 – 3 – половой процесс у базидиомицетов (2 – образование дикариофитной гифы; 3 – схема деления дикариофитной гифы и образование базидиоспор).
У гомоталличных (обоеполых) видов мужские и женские ядра находятся в разных клетках одного и того же мицелия. Образование двуядерных клеток происходит путем перехода мужского ядра в клетку, содержащую женское ядро. Такой диплоидный мицелий служит для формирования телиоспор головневых (класс Ustomycetes) и ржавчинных грибов (класс Teliomycetes). Переход от гаплоидного состояния к дикариотическому при объединении ядер клеток мицелиев, которые одинаковы морфологически, но различны в половом отношении, называют гетероталлизмом (греч. heteros – различный + thallos – тело). У гетероталличных видов в большинстве случаев происходит слияние и срастание двух одноядерных мицелиев, несущих мужские и женские ядра. На двуядерном мицелии чаще образуются плодовые тела или непосредственно на мицелии формируются базидии (отдел Basidiomycоtа). В дальнейшем в гимениальном слое плодовых тел из телиоспор головневых и ржавчинных грибов и непосредственно на двуядерном мицелии формируются базидии. Базидия представляет собой клетку булавовидной или цилиндрической формы, на поверхности которой формируются стерильные выросты – стеригмы. На стеригмах экзогенно образуется четное количество (чаще четыре) базидиоспор. Развитие базидии начинается с диплоидной клетки, которая находится на конце гифы. Образовавшееся в результате копуляции ядро проходит редукционное и миотическое деление, во время которого образуется четыре гаплоидных ядра, переходящих в базидиоспоры. Различают два основных типа базидий: – холобазидии – одноклеточные. Они образуются на конечных разветвлениях эндофитного мицелия, на гименофоре плодовых тел (различных по форме, величине и строению); – гетеробазидии (фрагмобазидии) – многоклеточные, имеющие продольные и поперечные перегородки. Они формируются из телиоспор ржавчинных и головневых грибов после их прорастания. Базидиоспоры после прорастания дают недолговечный гаплоидный мицелий. Между гаплоидными гифами гомоталличного или гетероталличного мицелия с помощью анастомозов вновь образуется дикариотичный мицелий, и цикл гриба повторяется. Способность одного и того же гриба давать несколько типов различных по форме, происхождению и функциям спороношений называется плеоморфизмом. Последовательное прохождение этих спороношений одного за другим в строго определенном порядке, завершающееся образованием исходных спор, называется циклом развития гриба.
Рисунок 9 – Способы формирования базидий с базидиоспорами: 1 – на конечных разветвлениях эндофитного мицелия (род Exobasidium); 2 – на гименофоре плодовых тел; 3 – из телиоспор головневых (а – холобазидия, род Tilletia; б – гетеробазидия, род Urocyctis), 4 – на ржавчинных грибах (гетеробазидия, род Phragmidium).
Грибы представляют собой очень обширную группу (тип) растений, содержащую до 80 000 описанных видов. Возможно, впрочем, что некоторое количество из указанного числа представляет одни и те же виды, но повторно описанные под разными названиями. Однако, с другой стороны, нужно признать, что видовой состав грибов изучен еще далеко не полно. Особенно это относится к тропическим областям, где, вероятно, будет найдено еще огромное количество новых видов. Поэтому некоторыми микологами-систематиками и флористами высказывается мнение, что грибы по общему числу видов, существующих сейчас на земле, мало чем уступают, а может быть, даже превосходят цветковые растения (около 150 000 видов). При указанном богатстве видами грибы отличаются вместе с тем большим разнообразием их. Стоит только вспомнить, что сюда относятся столь различные представители, как всем известные шляпные почвенные грибы, или древесные трутовики с их крупными сложно построенными плодовыми телами, далее — крупные, иногда огромные дождевики (до 1,5 м в диаметре у американской Calvatia) затем различные подземные клубневидные грибы, как трюфели и др. С другой стороны, к грибам относится еще значительно большее число микроскопических форм, заметных невооруженному глазу в виде различных налетов и плесеней на тех или других субстратах, а нередко и совсем неразличимых без микроскопа.
При всем этом разнообразии можно указать ряд черт, свойственных всем или огромному большинству грибов. Во-первых, все без исключения грибы не содержат хлорофилла и поэтому имеют гетеротрофное питание, т. е. черпают свой углерод только из готовых органических соединений. В соответствии с этим стоит строение их вегетативного тела. За исключением небольшого числа наиболее примитивных видов, живущих как внутриклетные паразиты зеленых растений, почти у всех остальных оно развито в виде мицелия, или грибницы, и представляет систему очень тонких ветвящихся нитей, или гиф, непрерывно нарастающих своими концами и пронизывающих субстрат, на котором развивается гриб. Благодаря такому строению мицелия, он имеет очень большую поверхность, через которую осмотическим путем и поступают в него питательные вещества, в том числе и сравнительно мало подвижные, но необходимые для него органические вещества.
При микроскопическом изучении обнаруживается, что мицелий различно построен у низших и высших грибов. У первых слагающие его гифы, хотя и достигают многих сантиметров в длину, но лишены поперечных перегородок, и, таким образом, весь мицелий оказывается, формально говоря, одной сильно разветвленной клеткой с огромным числом ядер в ней. Такой мицелий называется одноклеточным, или не членистым. У всех высших грибов и немногих низших гифы мицелия перегорожены поперечными перегородками на отдельные клетки, каждая с одним или чаще с несколькими ядрами. Такой мицелий называется многоклеточным, или членистым. За этим, хотя и существенным различием, мицелий всех грибов построен весьма сходно и одинаково пронизывает субстрат или (при большой влажности атмосферы) отчасти растет сверх него, образуя пушистый налет или более объемистые ватообразные скопления большею частью беловатого цвета.
Мицелий построен в общем слишком однообразно, чтобы по нему можно было идентифицировать тот или другой гриб. Более разнообразны и характерны для того или иного вида или группы видов органы размножения, приносящие споры и называемые поэтому также органами спороношения. При этом здесь различают бесполые и половые спороношения. Первые, подобно вегетативному размножению высших растений, служат для быстрого размножения и повторяют, как правило, полностью особенности произведшего их организма. Вторые имеют в основе половой процесс, сопровождающийся слиянием клеточных ядер и следующим затем редукционным делением их. Поэтому споры полового размножения, подобно семенам высших растений, не являются все вполне тождественными генетически друг с другом и с материнским растением.
Половые споры нередко представляют покоящиеся споры и служат в этом случае не столько для непосредственного размножения, сколько для сохранения вида в течение неблагоприятного периода (зима, засуха).
Среди бесполых спор различают эндогенные и экзогенные. Первые имеются только у низших грибов и образуются в большом числе внутри особых клеток, называемых спорангиями, почему и самые споры называют спорангиоспорами. Экзогенные споры, иначе называемые конидиями, свойственны высшим грибам и только немногим низшим. Они образуются не внутри, а на поверхности образующего их органа, называемого конидиеносцем.
Что касается половых спор, то они также бывают различны. У низших грибов они имеют характер покоящихся спор и различаются по форме полового процесса, давшего им начало (ооспоры, зигоспоры). При прорастании их развивается спорангий со спорами в нем, происшедшими в результате редукционного деления (т. е. генетически не однородными). У высших грибов половой цикл сложнее. Продукт оплодотворения у них не превращается в покоящуюся спору, а разрастается далее и в конце своего развития приносит многочисленные спороносные органы. Одни из них образуют внутри эндогенные споры и называются сумками, или асками, а их споры — аскоспорами. Другие образуют на своей поверхности экзогенные споры и называются базидиями, а их споры — базидиоспорами. Как аскоспоры, так и базидиоспоры образуются в результате редукционного деления, т. е. являются генетически неоднородными, как это свойственно вообще спорам полового происхождения (см. ниже споры спорангиев, получающихся при прорастании половых спор низших грибов).
Для пояснения вышеизложенного можно рассмотреть несколько конкретных примеров.
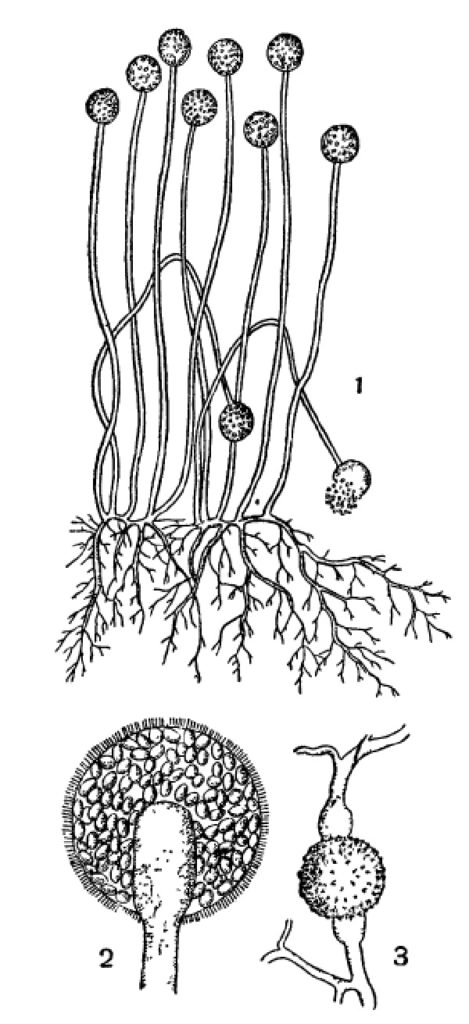
Обычнейший плесеневый гриб, Mucor mucedo, принадлежит к низшим грибам. Его бесполые спорангиоспоры выпадают из спорангия и, перенесенные токами воздуха на субстрат подходящего состава и влажности, немедленно прорастают. При этом спора вздувается и затем из нее вырастают 2—3 ростковые трубочки, которые быстро растут своими концами дальше, ветвятся и развиваются в нечленистый (одноклеточный или, лучше, неклеточный) мицелий (рис. 1). Он отчасти пронизывает субстрат, отчасти стелется по его поверхности. Через 2—3 дня роста на таком мицелии развиваются особые ветви, которые не ветвятся и растут прямо вверх на высоту до 10 см. Они представляют спорангиеносцы. Их концы шаровидно вздуваются, отделяются от спорангиеносца перегородкой и превращаются в спорангии (рис. 1). В спорангиях без всякого редукционного деления образуются спорангиоспоры таким образом, что общее протоплазменное содержимое распадается на большое число участков, одевающихся каждый собственной оболочкой (рис. 1, 2). Спорангиоспоры, выпадая вследствие разрушения оболочки спорангия, немедленно могут прорастать в такой же мицелий с новыми спорангиями на нем и т. д.
Рис. 1. Mucor mucedo. 1 — спорангиеносцы со спорангиями и мицелий; 2 — спорангий, сильно увеличенный; 3 — зигоспора на месте слияния двух одинаковых ветвей мицелия.
У Mucor mucedo имеется и половой процесс. Он происходит лишь при определенных условиях: при встрече двух различных в половом отношении мицелиев. При этом специальные короткие ветви гиф того и другого сливаются «своими концами, и из этого слияния получается зигоспора, одевающаяся толстыми оболочками и переходящая в состояние покоя (рис. 1,3). Позднее она прорастает в короткую нить с спорангием на конце. В нем на этот раз происходит редукционное деление при формировании спорангиоспор, так что последние в этом так называемом зародышевом спорангии оказываются генетически разнородными, отличаясь, между прочим, половыми свойствами развивающихся из них мицелиев.
Другой также очень обычный плесеневый гриб, Aspergillus, принадлежит к высшим грибам и имеет членистый многоклеточный мицелий. На нем развиваются конидиеносцы в виде вертикально растущих ветвей длиной в 1— 2 мм, вздувающихся на конце. На поверхности этого вздутия образуются короткие цилиндрические выросты, отчленяющие на своих концах последовательно целые цепочки конидий (рис. 2). Последние отпадают и, попав на подходящий субстрат, легко прорастают, давая 2—3 ростковые трубочки, нарастающие концами и ветвящиеся. В отличие от Mucor, они вскоре делятся поперечными перегородками, так что получается многоклеточный мицелий. На нем опять вскоре формируются такие же конидиеносцы и т. д.
Рис. 2. Aspergillus. 1 — конидиеносец с цепочками конидий; 2 — половые органы; 3 — развитие плодового тела (спиральное тело, в середине — разросшийся женский половой орган с зачатками аскогенных гиф на нем); 4 — часть разреза через аскокарп с сумками и аскоспорами в них.
На том же мицелии развивается и половое спороношение. Вначале появляются половые органы в виде парами расположенных коротких ветвей, часто одной и той же гифы. Одна из этих ветвей, спирально закрученная, является женским органом, а другая, более прямая — мужским (рис. 2, 2). Вскоре затем женский орган дает боковые короткие выросты, называемые аскогеными гифами. На их концах формируются сумки, а в них сначала происходит слияние двух имеющихся ядер и затем редукционное деление копуляциояного ядра. После следующих затем еще двух делений в сумке оказывается восемь ядер, и около них в каждой сумке формируется по восемь аскоспор. Последние, как происходящие в результате редукционного деления, оказываются такими же неоднократными в генетическом отношении, как и споры Mucor в зародышевом спорангии.
Одновременно с указанным развитием половых органов происходит обрастание их соседними ветвями мицелия, которые плотно сплетаются в округлое тельце, и уже внутри него развиваются аскогенные гифы и сумки. Аскоспоры освобождаются вследствие разрушения оболочки этого тельца, называемого сумчатым плодовым телом, или аскокарпом (рис. 2, 4).
Третим и последним примером можно взять какой-нибудь из шляпных грибов, например часто развивающийся на навозе Coprinus (рис. 3). На его многоклетном мицелии могут развиваться, хотя и не особенно обильно, бесполые (конидиальные) спороношения, а затем залагаются плодовые тела, или базидиокарпы, состоящие из пенька и шляпки. На нижней стороне последней развиваются в очень большом числе спороносные органы — базидии. Так же как и сумки, они имеют сначала по два ядра, которые затем сливаются, и после этого копуляционное ядро делится редукционным путем. Затем следует еще одно деление, и в базидии получается четыре ядра. Тем временем на ее поверхности образуются четыре выроста, и в них переходит по одному ядру (рис. 3, 2). Таким образом, на каждой базидии образуется по четыре экзогенных споры, называемых базидиоспорами. По характеру своего развития, при котором происходит слияние клеточных ядер и затем редукционное деление, они сходны с аскоспорами и должны быть отнесены к половым спороношениям. Как те, так и другие, в отличие от бесполых спор, оказываются генетически неоднородными.
Риге. 3. Coprinus. 1 — плодовое тело с остатками диплоидного мицелия у основания ножки; 2 — базидии (b) с 4 базидиоспорами каждая.
Полиморфизм.
Из всех указанных органов — мицелия и органов спороношения — первый построен настолько однообразно, что идентификация гриба по нему большей частью совершенно невозможна. Характерные признаки приходится поэтому искать во вторых. Однако один и тот же гриб может иметь их несколько: половые спороношения и бесполые, притом последних нередко целый ряд (до 5—6) и иногда весьма различных. Благодаря этому гриб может в корне менять свою физиономию на разных стадиях, так что, если бы не знать генетической связи между ними, то их пришлось бы принять за совершенно разные организмы. Раньше обыкновенно так и делалось. Например, конидиальные спороношения разобранного выше в качестве примера Aspergillus назывались Aspergillus glaucus, а сумчатые — Eurotium herbariorum. Также для представителей современного рода ржавчинных грибов, Puccinia, одно спороношение носило название Spermogonium, другое — Aecidium, третье — Uredo, четвертое — собственно Puccinia, — и все это не в качестве стадий развития одного и того же организма, а в качестве самостоятельных даже родов. Позднее, при изучении полного цикла развития и установлении генетической связи спороношений, эти названия утратили прежний смысл и, если сохранились иногда, то лишь для обозначения того или иного спороношения (например, в приведенном выше примере ржавчинных грибов — названия Aecidium, Uredo и др.).
Это явление — способность грибов менять свой внешний вид в состоянии разных спороношений — получило название полиморфизма, или плеоморфизма их.
Значение для идентификации грибов разных спороношений.
Ввиду указанного полиморфизма встает вопрос, насколько то или иное спороношение может характеризовать данный гриб. Здесь приходится различать органы бесполого и полового спороношений. Поскольку дело касается первых (бесполых), то в некоторых случаях они вполне типичны для данного круга форм и не повторяются у других. Таковы, например, спорангии у мукоровых.
То же, пожалуй, хотя и не с такой точностью, можно сказать относительно конидиеносцев Aspergillus и некоторых других. Однако очень часто бесполые спороношения не достаточно характерны для установления систематического положения гриба. Таковы, например, конидиеносцы так называемого типа Oedocephalum, похожие отчасти на Aspergillus, но отличающиеся тем, что конидии сидят на вздутом кончике конидиеносца не цепочками, а в один только этаж. Они встречаются у Peziza vesiculosa из аскомицетов, у Fomes annosus из базидиомицетов (трутовиков), у Cunninghamella из мукоровых, у Basidiophora из пероноспоровых, а, кроме того, существует еще немало форм с такими же конидиеносцами, у которых неизвестны, кроме них, иные спороношения и которые причисляются поэтому к группе несовершенных грибов (Fungi imperfecti) (рис. 4, 2). Другой пример: похожие на Oedocephalum, но не вздутые на конце конидиеносцы типа Botrytis известны у многих несовершенных грибов (не имеющих, кроме них, других спороношений); кроме того, они развиваются как бесполое спороношение у сумчатого гриба Sclerotinia fuckeliana (рис. 4, 2), тогда как у многих других видов того же рода Sclerotinia бесполые спороношения совсем иного рода, в виде так называемой Моnilia (цепочка конидий на коротком конидиеносце). Если бы основываться здесь на бесполых спороношениях, то пришлось бы разбить очень характерный и хорошо отграниченый род Sclerotinia. Наоборот, половые спороношения оказываются очень характерными не только для идентификации данного гриба, но и для установления его филогенетических отношений. Поэтому они кладутся в основу изучения, и на основании их производятся систематические группировки.
Рис. 4. Конидиеносцы: 1 — типа Oedocephalum; 2 — типа Botrytis.
Половое воспроизведение.
Ввиду того, что половое воспроизведение грибов весьма своеобразно и у многих изменило свою первоначальную форму, для понимания необходимо вкратце остановиться здесь на его цитологической основе. Типичный половой процесс в основе своей имеет слияние двух клеток (гамет) и их клеточных ядер. Получившееся при этом копуляционное ядро содержит двойное число хромосом и называется диплоидным. В известный момент развития, разный для разных организмов, диплоидное ядро претерпевает редукционное деление, которым снова восстанавливается гаплоидное число хромосом. Таким образом, для каждого организма, имеющего половой процесс, характерно, что в его цикле развития имеется чередование гаплоидного и диплоидного состояний, или ядерных фаз (от копуляции до редукции — диплоидная фаза, от редукции до новой копуляции — гаплоидная).
Если подойти с этой точки зрения к половому воспроизведению грибов, то можно заметить следующее: у низших грибов (архимицетов и фикомицитов) непосредственным результатом оплодотворения является зигота, имеющая характер покоящейся споры. При прорастании ее копуляционное диплоидное ядро (или ядра, если как у Mucor оплодотворение заключалось в слиянии двух многоядерных клеток) делится редукционным способом, благодаря чему восстанавливается гаплоидное число хромозом. Таким образом, здесь диплоидная фаза представлена только покоящейся зиготой, и вся активная жизнь гриба проходит в гаплоидном состоянии.
У аскомицетов мы также наблюдаем оплодотворение. Однако при этом за слиянием половых клеток, или цитогамией (вернее плазмогамией) не следует сейчас же слияния их ядер, или кариогамии. Мужские и женские ядра в оплодотворенном женском половом органе (архикарпе) только сближаются попарно друг с другом, образуя так называемые синкарионы, или дикарионы.
При разрастании половых органов и развитии из них аскогенных гиф дикарионы размножаются, причем в ряде случаев прослежено, что оба ядра делятся строго одновременно и параллельно друг другу. В молодой сумке происходит слияние этих двух ядер дикариона в одно копуляционное диплоидное ядро. Оно затем сейчас же претерпевает редукционное деление, так что аскоспоры содержат уже гаплоидные ядра и прорастают в гаплоидный мицелий. Часть этого цикла развития от плазмогамии до кариогамии может быть отождествлена с диплоидной фазой, так как каждый содержащийся здесь дикарион ведет себя как нечто единое и содержит двойное число хромосом. Поэтому по существу является второстепенным моментом, что они заключены здесь в две ядерные оболочки, а не в одну. Таким образом, у аскомицетов чередование ядерных фаз выражается в следующем: от аскоспор до плазмогамии идет гаплоидная фаза; от плазмогамии до кариогамии, в сумке — диплоидная фаза, или, как ее называют еще по характерному строению ядерного аппарата, дикариофит. В отличие от низших грибов диплоидная фаза представлена здесь не покоящимся состоянием (зигоспорой), а имеет известное разрастание и размножение своих ядер. Однако преобладание в цикле развития имеет все-таки гаплоидная фаза, так как на ее долю приходится не только вегетативный мицелий, могущий неограниченно размножаться путем образования бесполых спороношений, но и значительная часть сумчатого плодового тела (вся оболочка его и часть внутренней массы, кроме аскогенных гиф и молодых сумок). Нужно отметить еще, что у многих аскомицетов слияния половых клеток совсем не происходит, но всегда тем или иным образом (апогамия) происходит образование дикарионов и развитие аскогенных гиф, так что характер смены ядерных фаз при этом по существу не изменяется.
Что касается, наконец, базидиомицетов, то у них совершенно неизвестно образования половых клеток. Базидиоспоры здесь, как и аскоспоры, гаплоидны. При прорастании они дают гаплоидный, или первичный мицелий, который может размножаться бесполым путем, но в общем имеет непродолжительное существование и вскоре заменяется диплоидным, или вторичным мицелием. Последний возникает в результате анастомозов и слияний гиф одного первичного мицелия или чаще двух разных. Через анастомоз переходит ядро из одной клетки в другую, в результате чего получаются первые двуядерные клетки с дикарионами. Они затем разрастаются в целый мицелий, сложенный из таких двуядерных клеток (при этом, конечно, дикарионы размножаются подобно тому, как и в аскогенных гифах). Этот вторичный мицелий, относящийся к диплоидной фазе (дикариофит), обычно имеет более продолжительное существование и часто бывает даже многолетним. За счет его полностью слагается все плодовое тело. В развива юпщхся на нем базидиях, так же как и в сумках, происходит, наконец, слияние ядер дикариона, и копуляционное ядро сейчас же делится затем редукционным образом, так что развивающиеся на базидии базидиоспоры содержат уже по одному гаплоидному ядру. Базидиоспора снова прорастает в первичный гаплоидный мицелий. Таким образом, и у базидиомицетов мы имеем ясную смену ядерных фаз: от базидиоспоры до возникновения первых дикарионов — гаплоидная фаза; от первых дикарионов через весь вторичный мицеЛий и все плодовое тело до базидий — диплоидная фаза, цитологически представленная, как и у аскомицетов, в виде дикариофита, но в отличие от них здесь преобладающая. Из приведенного видно, что базидиальное спороношение, подобно сумчатому стоящее в связи со сменой ядерных фаз (рис 5), является по существу выражением половой функции. Поэтому оно вместе с половыми спороношениями других грибов привлекает особое внимание миколога, так как здесь находят наиболее надежные указания на филогенетические отношения и эволюцию изучаемых групп.
Однако было бы односторонним ограничиваться только статическим изучением половых спороношений. Истинная природа всякого организма лучше выясняется в его динамике, особенно в индивидуальном развитии, или онтогенезе. Особенное значение имеет такой подход для грибов с их часто очень сложным циклом развития. Поэтому современное морфолого-систематическое изучение грибов, дающее в первую очередь основу для понимания их истинной природы и филогенетических взаимоотношений, ставит перед собой более сложные задачи: выяснение полного цикла развития с учетом цитологических явлений и сравнительное изучение не только строения, но и развития как вегетативных органов, так и различных спороношений, образующихся в той или другой фазе развития.
Настоящее, второе, издание «Микологии» сохраняет тот же план и те же основные установки, как и первое, опубликованное в 1933 г. Как и раньше, в нем ставится задача дать некоторый синтез знаний о грибах, не ограничиваясь их морфологией и систематикой, но обращая также значительное внимание на условия их жизни, значение в природе и хозяйстве и в связи с этим на те экспериментальные направления, которые приобретают все большее значение в современных микологических исследованиях.
Автор попрежнему полагает, что такое построение более соответствует запросам, предъявляемым у нас к микологии, притом не только со стороны ботаников, но и со стороны фитопатологов, агрономов, техников и др., чем те очень хорошие, но только морфолого-систематические сводки, какие появились за последние годы в Западной Европе и Америке (Gaumann, Gwynne-Vaug- han, Bessey). В противоположность им можно указать на старое классическое руководство Де-Бари (Vergleichende Morphologie und Biologie der Pilze, 1884), которое еще более 50 лет назад широко трактовало микологию: не только как морфологию и систематику грибов, но и как их биологию, понимаемую в широком смысле и с особенным ударением на явлениях паразитизма.
Посвящая настоящее издание памяти А. Де-Бари в связи с исполнившимся в этом году пятидесятилетием со дня смерти этого величайшего исследователя- миколога, автор хотел вместе с тем указать, что современное положение нашей науки опять приводит нас к той же широкой трактовке ее, как это было заложено в его классических сочинениях.
Понятно, что фактический материал, каким мы сейчас располагаем, значительно обширнее и не может быть поэтому изложен с той почти исчерпывающей полнотой, как это было сделано Де-Бари в его руководстве 1884 г. Наша книга и не претендует на исчерпывающее изложение. В основном — это руководство, стремящееся отразить только основные факты и главнейшие направления в современной микологии, но вместе с тем оно имеет в виду читателя, предъявляющего более глубокие запросы к этой дисциплине. Предполагается, что это будут главным образом студенты старших курсов, специализирующиеся в этой области, аспиранты и начинающие научные работники, притом не только ботаники и фитопатологи, но отчасти и агрономы, техники и др.
Для некоторых из этих категорий предполагаемых читателей было бы, вероятно, желательно иметь также побольше справочного и методического материала. Однако автор не решился расширить книгу в этом направлении из опасения слишком большого увеличения ее объема. Тем более, что мы имеем уже сейчас очень основательные справочники Н. А. Наумова по методике культуры и микроскопического изучения грибов. Также обильный справочный материал по некоторым вопросам можно найти в «Основах микологии» А. А. Ячевского (1933). Эта книга, остановившаяся из-за смерти автора на первом томе, касается только некоторых разделов общей микологии, но благодаря обилию справочного материала, содержит более 1000 страниц. Можно думать, что обработка в том же масштабе всей микологии потребовала бы, вероятно, по крайней мере, еще двух таких же томов. В наши намерения совершенно не входило составление такого огромного руководства.
При сохранении плана и общих установок первого издания тем не менее второе издание значительно переработано и расширено. Это коснулось не только исправления исключительно обильных опечаток первого издания, печатавшегося очень спешно и во многих листах почти без корректуры, но и исправлений и дополнений по существу в связи с данными новейшей литературы и необходимостью отразить некоторые новые течения, недостаточно выявленные и даже совсем отсутствовавшие в то время, когда изготовлялась рукопись первого издания (была окончена и сдана в издательство в 1931 г.). Также были приняты во внимание некоторые критические замечания по поводу первого издания, полученные от товарищей по науке, за что автор выражает им свою признательность. Впрочем, не все из их пожеланий могли быть выполнены. Так, указывалось на желательность расширения отделов по систематике пиреномицетов и несовершенных грибов. Этого не сделано, по крайней мере, в значительных размерах. Вполне соглашаясь, что каждый миколог должен знать значительное число представителей этих крупнейших групп, автор тем не менее полагает, что такие знания должны получиться из гербаризации и определения, чего не может заменить учебное руководство. Сделать же какие- либо существенные обобщения на основании более подробной классификации пиреномицетов и несовершенных грибов автор не видел возможности.
Как наиболее существенные изменения второго издания можно указать следующие:
В целом несколько смягчено догматическое изложение различных гипотез и толкований (особенно в вопросах филогенеза) приведением других существующих мнений на этот счет и их кратким критическим разбором. Расширена глава о гетероталлизме. Расширено изложение учения о биотипах и их возможном гибридном происхождении. Последнее сделано как в общей главе о паразитизме, так и более специально в главе о ржавчинных, где эти явления наилучше изучены. Переработана глава VIII — об общих условиях жизни грибов—и расширена изложением учения о значении ростовых веществ и биоса, а также явлений антагонизма. Расширено изложение учения о симбиозе. Расширены в систематической части главы о головневых и особенно ржавчинных грибах. Приводимые в конце глав списки литературы значительно расширены. Однако и в таком виде они отнюдь не могут претендовать на исчерпывающую полноту. Заново написан довольно подробный очерк истории микологии. Он приложен в конце книги, так как построен таким образом, чтобы показать развитие тех проблем, современное состояние которых излагается в основной части.
В общем, в результате указанных и многих других изменений и расширений общий объем текста увеличился примерно на 20—25%, а число иллюстраций — с 237 до 326.
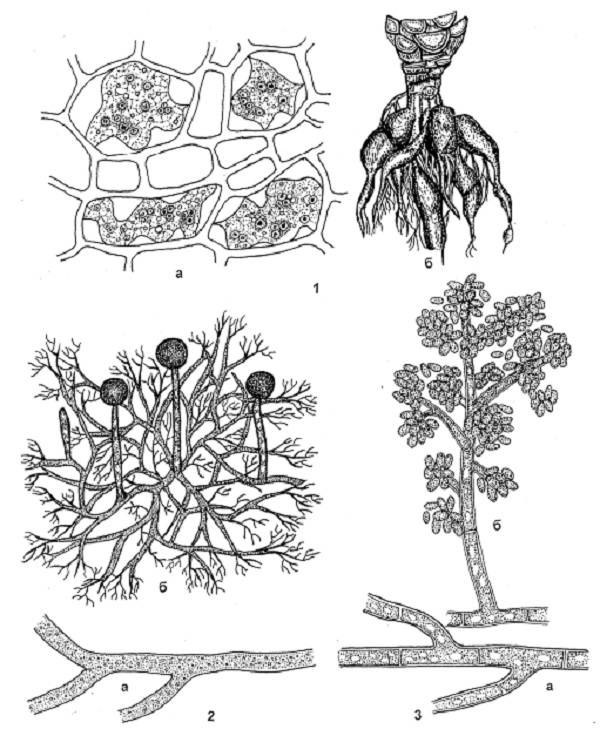
Вегетативное тело у наиболее примитивных грибоподобных протистов, псевдогрибов и настоящих грибов отдела Хитридиомицеты представлено плазмодием. Плазмодий, или амебоид – многоядерная, лишенная оболочки, изменчивой формы клетка миксомицетов, плазмодиофоровых и низших хитридиевых грибов. У большинства настоящих грибов вегетативное тело состоит из тонких ветвящихся нитей – гиф, имеющих вид цилиндрических трубок до 10 мкм в диаметре. Они характеризуются апикальным (верхушечным) ростом и обильным ветвлением. Совокупность гиф называется грибницей, или мицелием. Он может быть неклеточным, или несептированным (у низших) и многоклеточным, или септированным – у высших грибов. Клетка грибов, за исключением зооспор и вегетативного тела у некоторых низших, окружена твердой оболочкой. Состав веществ клеточной оболочки разнообразный. Так, в клеточных оболочках оомицетов преобладают целлюлозно-глюкановые полисахариды, у зигомицетов – целлюлозно-хитиновые, у хитридиевых, сумчатых, базидиальных и дейтеромицетов – хитино-глюкановые соединения. Обычно оболочки молодых клеток тонкие, бесцветные, однородные по структуре (рисунок 1). С возрастом оболочка может утолщаться, ослизняться, менять окраску в результате отложения пигмента. Через поверхность грибницы осмотическим путем происходит всасывание питательных веществ. Мицелий по отношению к растению – хозяину может быть поверхностным или внутренним. Если мицелий развивается на поверхности пораженного растения или другого питательного субстрата, его называют поверхностным, или экзогенным. Такой мицелий имеется, например, у возбудителей настоящей мучнистой росы.
Мицелий, распространяющийся внутри субстрата, называют внутренним, или эндогенным (греч. endon – внутри). В свою очередь эндогенный мицелий у некоторых представителей проникает в клетку растения-хозяина (Olpidium brassicaе Dang., Synchytrium endobioticum Pers.), но у подавляющего большинства грибов он распространяется по межклетникам (возбудители ложной мучнистой росы, ржавчинных заболеваний и др.).
Видоизменения мицелия грибов
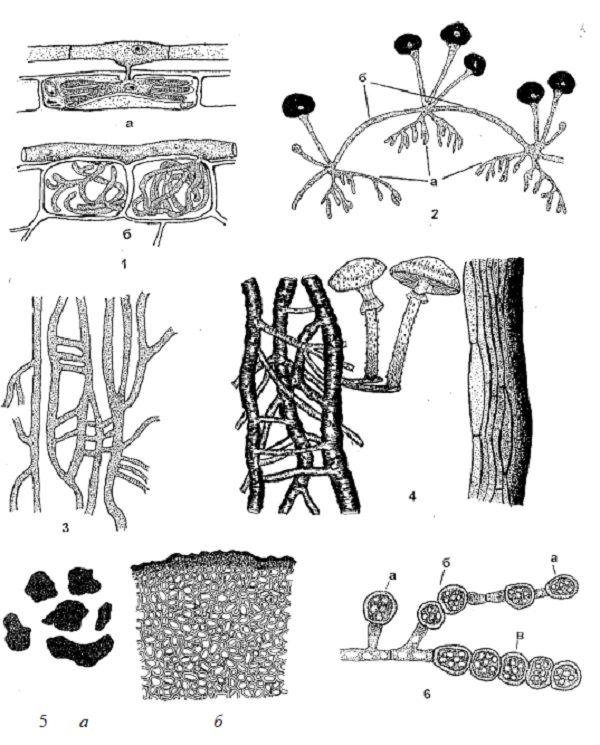
В зависимости от условий развития и выполняемых функций мицелий или отдельные гифы грибов могут видоизменяться (рисунки 2 и 3). Пряжки – полукруглые клетки, расположенные сбоку гиф в местах перегородок и связывающие полости соседних клеток. Пряжки характерны для многих базидиальных грибов. По ним при половом процессе перемещаются содержимое и ядро из одной клетки в другую. Наличие или отсутствие пряжек, их форма и количество являются систематическим признаком для гименомицетов. Анастомозы (греч. anastomosis – соединение) боковые короткие выросты клеток, соединяющие гифы мицелия между собой. По ним протоплазма и ядро из одной клетки могут переходить в другую. В отдельных случаях с помощью анастомозов осуществляется переход мицелия от гаплоидного состояния к диплоидному. При сильном развитии анастомозов мицелий становится сетчатым. Аппрессории (греч. apprimere – прижимать) – расширенные или лопастные выросты гиф, с помощью которых возбудители фитофторы, настоящей мучнистой росы прикрепляются к поверхности субстрата. Гаустории (лат. haustor – черпающий, пьющий) – боковые, лопастновидной или гифоподобной формы, выросты мицелия грибов, характерные для облигатных фитотрофов. Гаустории имеют булововидную форму. Они проникают в клетки растения-хозяина и транспортируют питательные вещества из клетки к мицелию. Ризоиды (греч. rhiza – корень + еidos – вид) – простые или разветвленные корневидные отростки гиф, с помощью которых гриб проникает в субстрат и прикрепляется к нему. Столоны (лат. stolo, stolonis – корневой побег) – дугообразные толстые гифы, с помощью которых гриб быстро распространяется на субстрате. Ризоиды и столоны есть у вида Rhizopus nigricans Ehr. – возбудителя черной головчатой плесени. Придатки (лат. appendix – придаток) – специальные, различной формы и размера, бесцветные или окрашенные гифы, одно- или многоклеточные. Они отрастают от оболочки клейстотециев и удерживают плодовые тела на поверхности субстрата, способствуя их распространению. Придатки всех видов мучнисторосяных грибов являются важным систематическим признаком при классификации этих грибов.
Рисунок 2 – Видоизменения мицелия грибов: 1 – гаустории у грибов рода Erysiphe (a) рода Peronospora (б); 2 – ризоиды (а) и столоны (б) у грибов рода Rhizopus; 3 – анастомозы; 4 – ризоморфы у грибов рода Armillariella; 5 – склероции у грибов рода Sclerotinia; 6 – хламидоспоры (а – терминальные, верхушечные; б – интеркалярные, промежуточные; в – в цепочках у грибов рода Verticillium).Рисунок 3 – Видоизменения мицелия грибов: 1 – пряжки на гифах грибов; 2– мицелиальные придатки на клейстотециях; 3 – «реснички» на конидиях; 4 – специальные стерильные клетки: парафизы (а), щетинки (б), цистиды (в); 5 – мицелиальные тяжи; 6 – мицелиальные пленки (домовый гриб).
Реснички – нитевидные бесцветные клетки, расположенные на конце конидии в количестве от 2 до 5 шт. Они характерны для грибов рода Pestalotia и выполняют функцию удерживания конидий на поверхности растения-хозяина. Специальные стерильные гифы – парафизы – образуются у сумчатых и базидиальных грибов и выполняют разделительную или защитную функцию. Парафизы – одно – или многоклеточные бесплодные гифы, расположенные между сумками или базидиями. Они предохраняют их от высыхания и механических повреждений. Цистида – это стерильный, обычно светлоокрашенный, различных форм конец несосудистой гифы в гимениальном слое базидиальных грибов; щетинки – крупные толстостенные темноокрашенные клетки с шипиками на концах, возвышающиеся над базидиями. Хламидоспоры (лат. chlamyda – верхнее шерстяное платье) – одна из форм видоизменения мицелия, образующаяся путем распадения мицелия или отдельных его участков на самостоятельные клетки, которые округляются и под прежней клеточной стенкой покрываются плотной, толстой пигментированной оболочкой. Зачастую оболочка темноокрашенная, покрытая шипиками, щетинками, бугорками или сеточкой. Хламидоспоры содержат значительные запасы питательных веществ, поэтому способны сохраняться до 10, а по некоторым данным – до 15 лет. Формируются у всех факультативных сапро – и фитотрофов при неблагоприятных условиях среды. Геммы (лат. gemma – резной камень) образуются так же, как и хламидоспоры, но отличаются разнообразием форм. Они встречаются у сумчатых, базидиальных и несовершенных грибов. Мицелиальные тяжи (шнуры) очень часто образуются у различных групп высших грибов. Они бывают простые и сложные, разной длины, толщины, цвета и консистенции. В простейшем случае тяжи состоят из небольшого количества параллельно растущих гиф. Последние или склеиваются друг с другом, или образуют более прочные соединения при помощи многочисленных коротких анастомозов у домового гриба. Выполняют функции сохранения, распространения грибов при вегетативном размножении и служат своеобразной проводящей системой для передвижения питательных веществ и воды к развивающимся плодовым телам. Мицелиальные пленки – плотные мицелиальные сплетения, развивающиеся на поверхности или внутри питательного субстрата. У древоразрушающих трутовиков они формируются в трещинах древесины пораженного дерева. Пленки обладают высокой прочностью, иногда достигая в толщину 10–15 мм. Из мицелиальных пленок развиваются плодовые тела, мицелий или от них отходят тяжи. Ризоморфы (греч. rhiza – корень + morphe – форма) – сложные шнуровидные темные или черные сплетения гиф, напоминающие по форме корни высших растений. Их длина может достигать нескольких метров при толщине в несколько миллиметров. Гифы, образующие ризоморфы, различаются по строению и выполняемым функциям. Наружная часть ризоморф сложена из темных толстостенных гиф, внутренняя – из бесцветных тонкостенных гиф и гиф типа сосудов, т. е.неветвящихся, без перегородок, с широкими просветами. Ризоморфы выполняют функции сохранения гриба при неблагоприятных условиях и передвижения питательных веществ. Типичные ризоморфы образует опенок (Armillariella mellea Karst.). Склероции (греч. skleros – твердый) – это мицелиальные образования твердой, плотной консистенции, округлой, удлиненно-овальной, плоской или неправильной формы, размером от десятых долей миллиметра (микросклероции) до 30 см. Склероции состоят из темноокрашенного поверхностного слоя, который включает 1–4 слоя толстостенных округлых, плотно соединенных между собой клеток (параплектенхимы) и белой внутренней части, сформированной из рыхлого сплетения удлиненных тонкостенных бесцветных гиф (прозоплектенхимы). Они содержат до 10% воды и около 30% жира. Склероции грибов легко переносят неблагоприятные условия среды и длительное время сохраняются. Из них может развиваться мицелий или различные органы спороношения. Склероции известны у многих грибов, главным образом из родов Sclerotinia, Verticillium, Clavicеps и другие.