В этом порядке известно более 300 видов почти исключительно сапрофитов. Немногие известные здесь паразиты паразитируют главным образом на других мукоровых. Многие мукоровые чрезвычайно широко распространены в качестве спутников человека в виде плесеней (головчатая плесень) на всевозможных, растительных преимущественно, субстратах: хлебе, вареньи, овощах, плодах и т. д. В естественной обстановке наиболее обычными местообитаниями их являются экскременты травоядных животных и почва (см. стр. Копрофильные грибы и Почвенные грибы). Культуру мукоровых грибов очень легко получить, поместив свежий конский навоз под колпаком на тарелке. Обыкновенно уже через 3—4 дня появляется обильная флора мукоровых. Редко мукоровые встречаются в богатых органическими веществами водоемах (сточные воды). Они представлены там обыкновенно в виде стерильных мицелиев и образуют органы размножения только в воздухе.
Мицелий мукоровых состоит большею частью из бесцветных, сильно разветвленных, довольно толстых гиф, конечные разветвления которых обычно утончаются. Более специализированные пучки таких тонких, сильно разветвленных гиф носят название ризоидов. Они особенно развиты у Rhizopus и Absidia на их так называемых столонах, в месте их соприкосновения с субстратом (см. стр. Микроскопическое строение мицелия и рис. 22). Нормально мицелий у мукоровых не имеет перегородок, однако у значительного количества представителей они появляются в более старых частях и особенно в спороносных органах, например у Sporodinia. У Mucor racemosus и некоторых других наблюдается развитие перегородок и в молодых частях мицелия при культуре его погруженным в жидкость при недостаточной аэрации. В дальнейшем по этим перегородкам мицелий распадается на отдельные членики, или клетки, которые мы должны назвать по происхождению оидиями. Они здесь имеют шаровидную форму и размножаются далее почкованием, образуя так называемые мукоровые дрожжи (см. стр. Почкующийся мицелий рис. 26, 2). При перенесении на поверхность твердого субстрата они снова вырастают в мицелий.
Что касается полового воспроизведения мукоровых грибов, то, как известно, здесь впервые было обнаружено явление гетероталлизма (Blakeslee, 1904). В настоящее время с этой стороны изучено более 80 видов, и из них приблизительно ¾ оказались гетероталличными и ¼ гомоталличными. У гетероталличных форм, кроме нормальных (+) и (—) мицелиев, наблюдаются еще иногда нейтральные, которые не дают обычно копуляции ни с (+), ни с (—). Выше было указано на экспериментальное получение таких нейтральных мицелиев у Phycomyces путем смешения протопластов того и другого пола. Однако, кроме таких гетерокариотических мицелиев, отсутствие половой реакции наблюдается у многих рас, повидимому, просто в результате ослабления половой функции без всякого смешения разнородного ядерного материала. Можно думать, что такое ослабление довольно обычно и что им в значительной степени объясняется сравнительная редкость полового процесса у некоторых мукоровых. По форме половой процесс у мукоровых грибов представляет типичную зигогамию (рис. 122).

2 — Phycomyces blakesleeanus: a — начало процесса, b — готовая зигота, окруженная шиловидными выростами суспензоров;
3 — Моrtierella candelabrum, в b зигота окружена обверткой из гиф;
4 — Zygorrhynchus heterogamus;
5 — Mucor tenuis, азиготы;
6 — Piptocephalis freseniana;
7 — Absidia spinosa, выросты только на более крупном женском суспензоре (7, с).
Копуляция происходит между двумя укороченными и расширенными на концах ветвями, принадлежащими одному мицелию (у гомоталличных форм) или разным (у гетероталличных форм). Эти ветви на концах отделяют по многоядерной клетке, слиянием которых и формируется собственно зигота. В готовом состоянии она одета толстой бугристой или шиповатой оболочкой, один или два слоя которой окрашены в темнобурый, иногда почти черный цвет. Несущие ее копуляционные ветви называются подвесками, или суспензорами (рис. 122, 1). У некоторых форм, как у Phycomyces или Absidia, из них развиваются особые шиловидные выросты, отчасти охватывающие зиготу (рис. 122, 2, 7). У Mortierella аналогичные выросты суспензоров вместе с ветвями прилежащих гиф образуют плотную многослойную обвертку около зиготы, так что в результате получается нечто вроде плодового тела (рис. 122, 3). Копулирующие ветви и гаметангии на них у большинства представителей совершенно одинаковы, но у некоторых они показывают известные различия как в величине, так и в строении. Из гомоталличных форм такое различие в величине постоянно наблюдается, например, у представителей рода Zygorrhynchus (рис. 122, 4). У гетероталличной Absidia spinosa сюда добавляется еще то, что только один более крупный суспензор несет выросты, окружающие зиготу, тогда как другой остается гладким (рис. 122, 7). Такой же переход от типичной изогамии к гетерогамии намечается иногда по месту положения зиготы. У большинства форм она образуется прямо на месте двух слившихся клеток и лежит поэтому как раз между суспензорами; у других же она развивается из особого выроста, который у Piptocephalis, например, или у Pilobolus выходит как раз из средины копуляционной клетки (рис. 122, 6), но у Syncephalis nodosa, где гаметангии очень вытянутые и спирально обвиваются друг около друга, зигота образуется в качестве выроста одного из них на довольно заметном расстоянии от места их слияния. В этом и других подобных случаях гетерогамии гаметангии, несущий зиготу, можно назвать женским в отличие от другого — мужского.
Что касается внутренних процессов, происходящих при копуляции, то здесь прежде всего надо отметить, что сливающиеся в половом процессе клетки всегда многоядерны. Они соответствуют не гаметам, а целым гаметангиям, и за их слиянием, вероятно, следует множественная кариогамия. В большинстве изученных случаев число ядер в каждом гаметангии весьма значительно, например у Sporodinia grandis больше 1000, и все они (или почти все) копулируют друг с другом. Однако у некоторых представителей наблюдается, что копулируют только некоторые ядра, а остальные отмирают. У Zygorhynchus Dangeardii наблюдается всего только два диплоидных копуляционных ядра в зиготе. Самые слияния ядер здесь часто весьма запаздывают, а у Phycomyces blakesleeanus ядра сливаются только при прорастании и даже, может быть, только в зародышевом спорангии. Зиготы прорастают только после периода покоя: при этом они набухают, наружные окрашенные слои оболочек лопаются, и из внутренних частей вырастает слабо ветвящаяся или совсем неветвящаяся нить, которая заканчивается зародышевым спорангием (рис. 34, 4). Судя по тому, что образующиеся в нем споры оказываются неодинаковыми в генетическом отношении, надо полагать, что здесь происходит редукционное деление копуляционных ядер, перешедших сюда из зиготы (или, может быть, только одного из них; см. стр. Цитология и наследственность).
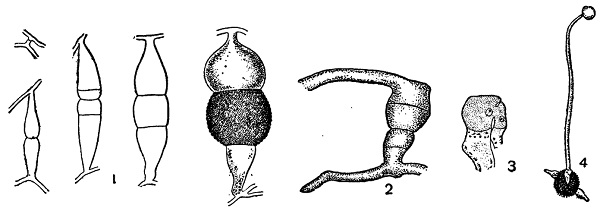
1 — Rhizopus nigricans;
2 — Absidia spinosa — гетерогамная копуляция;
3 — Endogone — гетерогамная копуляция одноядерных клеток;
4 — Мисоr — прорастание зигоспоры образованием зародышевого спорангия.
У некоторых мукоровых наряду с нормальными зигоспорами образуются еще азиготы из одного только гаметангия. Появление их зависит иногда от внешних условий, например излишне высокой температуры, а также обычна при попытках к гибридному оплодотворению. Однако в опытах Burgeff и особенно Saito и Naganishi удалось получить гибридные зиготы в ряде комбинаций (больше 10). Наконец, некоторые виды (например Mucor tenuis и некоторые другие), повидимому, совсем утратили половой процесс и образуют только азиготы (рис. 122, 5). Цитологические процессы при образовании азигот, равно как и их прорастание, до сих пор неизвестны.
Органы бесполого размножения у мукоровых в наиболее типичных случаях представлены спорангиями. Они образуются только вне субстрата в воздухе на концах особых спорангиеносцев, или прямо растущих кверху, или приподнимающихся (см. стр. Бесполое размножение. Споры → тропизмы органов спороношения). При развитии спорангия кончик спорангиеносца шаровидно вздувается, и затем у основания его залагается перегородка, идущая большею частью не прямо поперек, а изгибаясь куполообразно кверху по границе между более густой протоплазмой, собранной в верхней части вздутия (споронлазмой), и вакуолистым нижним участком. Эта куполовидная перегородка очерчивает так называемую колонку, представляющую как бы продолжение спорангиеносца, вдающееся в полость спорангия (рис. 123, 2). Протоплазма, содержащаяся в спорангии, распадается затем на отдельные споры, число которых может достигать тысяч и даже десятков тысяч на один спорангий (у Phycomyces — около 70 тысяч). К этому времени оболочка спорангия претерпевает изменения своих физических и химических свойств. Первоначально в молодом спорангии она имеет такой же в общем характер, как и оболочка спорангиеносца или мицелия, и обладает достаточной прочностью. Затем по мере созревания в ней происходят изменения состава, по некоторым указаниям — превращение в каллезу, в результате чего оболочка теряет свою прочность; одновременно с этим на ее поверхности часто отлагается оксалат кальция в виде игольчатых кристаллов, придающих спорангию как бы тонко шиповатую структуру. Оболочка зрелого спорангия у многих форм расплывается от воды и даже от влажного воздуха (например от дыхания), и споры при этом освобождаются (рис. 123, 2). Если это происходит при большой влажности, то находящаяся между спорами слизь (ее происхождение не ясно) взбухает и раздвигает споры друг от друга и вместе с тем выдвигает их из спорангия.

1 — зрелый спорангий с колонкой;
2 — выпадение спор.
Указанное расплывание оболочки наблюдается не у всех видов. Даже там, где оно хорошо выражено (например у Mucor mucedo), обыкновенно самая нижняя часть оболочки сохраняет большую прочность и остается в виде воротничка у основания колонки. В других случаях, например у Mucor racemosus, оболочка спорангия не расплывается, а разрывается на кусочки. Наконец, у некоторых, как у Pilobolus, оболочка спорангия, повидимому, сильно кутинизирована и отличается значительной прочностью. Спорангий здесь не раскрывается, а в замкнутом виде отбрасывается от спорангиеносца развиваемым в нем тургорным давлением (рис. 60). Что касается самых спорангиоспор, то они круглые или овальные, большею частью бесцветные, иногда коричневатые или дымчатые. Оболочка их гладкая, реже имеет скульптурную поверхность (например исчерченность у Rhizopus).

1 — общий вид при слабом увеличении;
2 — отдельный зрелый спорангиеносец;
3 — «выстрел».
Значительный интерес представляет развитие спор в спорангии. Последний с самого начала содержит большое число клеточных ядер. В дальнейшем, когда протоплазма спорангия начинает распадаться на споры, процесс этот протекает неодинаково у разных представителей. У большинства исследованных форм протоплазма спорангия прямо распадается на многоядерные участки, которые затем округляются, одеваются оболочкой и превращаются в споры (рис. 124).

У Circinella происходит распадение протоплазмы на одноядерные участки, так называемые протоспоры. Затем они еще разрастаются, делаются многоядерными и лишь тогда одеваются оболочкой, превращаясь в настоящие споры. У Pilobolus также залагаются сначала одноядерные протоспоры, но затем они не только разрастаются, но и делятся на несколько многоядерных участков, формирующихся в споры. Указанные различия, может быть, не лишены значения с точки зрения однородности или разнородности ядерного аппарата в спорах. Всюду они оказываются многоядерными, но, тогда как у Circinella и Pilobolus ядра их в недалеком прошлом сводятся на одно ядро протоспоры и потому могут рассматриваться как достаточно однородные, у остальных форм ядра в споре могут быть столь же разнородными, как во всем спорангии. Интересно, что в зародышевом спорангии, вырастающем из зиготы, развитие спор, поскольку это изучено до сих пор (у Phycomyces и Mucor), происходит несколько иначе. Именно, здесь споры залагаются одноядерными, аналогично протоспорам Circinella, но затем делаются многоядерными. Таким развитием особенно обеспечивается однородность ядерного аппарата в спорах зародышевого спорангия и, стало быть, их чистота в генетическом отношении. Поскольку дело касается наилучше изученного распределения половых признаков в самом деле споры зародышевого спорангия почти всегда оказываются «чистыми», или (+) или (—); лишь очень редко попадаются нейтральные, происходящие, вероятно, из случайно двуядерного зачатка, притом с неодинаковыми ядрами (см. стр. Цитология и наследственность).
У целого ряда мукоровых вместо спорангиев, содержащих значительное количество спор в каждом, развиваются на сильно разветвленных спороносцах отдельные конидии. Этим понижается энергия размножения, но вместе с тем, может быть, лучше обеспечивается развитие в сухопутных условиях, так как конидия формируется весьма быстро и не требует, как спорангий, значительного времени (сутки и больше) для полного развития. В эволюции мукоровых можно проследить, как спорангии превращаются в конидии, причем это превращение идет несколькими путями. В группе, объединяемой в семейство Chaetocladiaceae, это превращение происходит путем уменьшения спорангия до односпорового образования. Здесь у Thamnidium, весьма обычного сапрофита на навозе, имеется два сорта спорангиев. Один, сидящий на конце главной оси спорангиеносца, и другие, развивающиеся на ее боковых коротких ветвях. Первый спорангий — крупный, многоспоровый, с колонкой. Боковые спорангии, иначе называемые спорангиолями, — мелки, содержат около 10 и меньше спор и не имеют колонки (рис. 126, 1).

1 — Thamnidium elegans:
a — характер ветвления,
b — при большом увеличении конечный спорангий и боковые спорангиоли,
с — опавшие спорангиоли;
2 — Chaetocladium Brefeldii — конидиеносец с конидиями.
При неблагоприятных условиях главный спорангий совсем не развивается, а спорангиоли иногда уменьшают число спор до одной, т. е. биологически превращаются в конидию, хотя спора и не срастается здесь с оболочкой спорангиоля. Еще дальше дело пошло у Ghaetocladium, часто встречающегося паразита на других мукоровых. Здесь на сильно разветвленных спороносных гифах образуются исключительно односпоровые спорангиоли, причем споры срастаются с оболочкой спорангиоля и сваливаются вместе с ней, т. е. превращаются в действительные конидии (рис. 126,2). У Ch. Jonesii происхождение споры сказывается в момент ее прорастания, когда оболочка спарангиоля отделяется в виде экзоспория. У другого вида, Ch. Brefeldii, незаметно и этой двойственной оболочки. B другом круге форм, объединяемом в семейство Choanephorасеае, спорангий заменяется конидиями в результате перенесения развития спор из внутренних его частей на поверхность. У Choanephora могут, развиваться спорангии нормального строения с колонкой, и наряду с этим, при более обильном питании, на поверхности вздутого на конце спорангиеносца (который при этом не отделяется перегородкой), появляются выпуклины, которые и развиваются в экзогенные споры (рис. 127, 2).

1 — Choanephora cucurbit arum:
a — конидиеносец,
b — спорангий;
2 — Cunningham ella echinulata, конидиеносец (типа Oedocephalum);
3 — Blakeslea trispora:
a — нормальный спорангий,
b — экзогенные спорангии.
У Cunninghamella (рис. 127, 2) имеется только это второе спороношение, причем начальные стадии в виде вздувающегося на конце спороносца совершенно соответствуют развитию спорангия (но не отделяются перегородкой). У Blakeslea имеются и нормальные спорангии и превращение их в экзогенные спороношения. В этом последнем случае на поверхности основного вздутия, соответствующего зачатку спорангия, образуются выпуклинь. Последние, однако, развиваются не в конидии, а в небольшие трехспоровые спорангиоли, которые сваливаются и распространяются целиком как одна спора (рис. 127,3). Наконец, в семействе Piptocepbalidaceae (Syneephalastrum, Syncephalis и Piptocephalis) типичных спорангиев не развивается, но на поверхности зачатка спороношения формируются частичные спорангии или спорангиоли цилиндрической формы, и в них развиваются споры, расположенные в один ряд (рис. 128).

1 — Syncephalis:
а — спорангиеносец,
b — спорангий;
2 — Syncephalastrum, заложение экзогенных спорангиев;
3 — Piptocephalis:
a — спорангиеносец,
b — деталь одной ветви.
У Syneephalastrum эти споры большею частью одноядерны и лежат свободно в оболочке спорангиоля. У Syncephalis и Piptocephalis они срастаются с ней боковыми стенками и цилиндрический спорангиоль распадается при созревании на ряд одноклетных члеников, соответствующих каждый одной споре (т. е. здесь спорангиоль как бы превращается биологически в цепочку конидий).
Представляя одну из распространеннейших групп грибов, мукоровые, хотя и являются в большинстве случаев безразличными сапрофитами, тем не менее играют немалую роль как в природе, так и в жизни человека. Выше было указано на их большое количество в почве и значение в почвенных процессах (см. стр. Почвенные грибы). Специально в связи с деятельностью человека можно указать на значение мукоровых как плесневых грибов. Здесь как чистые сапрофиты особенно часто встречаются представители рода Мисоr и Rhizopus nigricans. Последний, впрочем, нередок и в полупаразитных условиях, на свежих фруктах и т. п. живых продуктах. У близкого Rh. tritici эта способность к паразитному развитию выражена еще сильнее, и он является, особенно в Америке, серьезным вредителем зерен злаков при недостаточно сухом хранении. Из паразитов на животных следует отметить Mucor corymbifer, Absidia Lichteimii, Мисоr pusillus, вызывающие иногда опасные поражения внутренних органов у человека. Часто также они поражают птиц.
Наконец, следует указать на бродильные свойства мукоровых. Способность производить алкогольное брожение в сахаристых средах присуща весьма многим формам, хотя и выражена часто не сильно; однако некоторые являются энергичными бродителями, мало даже уступая в этом отношении дрожжам. Весьма резко бродильная способность выражена у Mucor racemosus, M. genevensis, M. javanicus, M. rouxianus и некоторых других, причем некоторые, как М. racemosus и М. genevensis, бродят преимущественно в состоянии мукоровых дрожжей, а другие — в виде нормального мицелия. М. rouxianus и М. javanicus применяются на юго-востоке Азии при приготовлении алкогольных напитков (восточно-азиатские дрожжи). Они имеют даже известное преимущество перед настоящими дрожжами, так как сами осахаривают крахмал и применение их не требует поэтому предварительной обработки крахмалистых исходных продуктов. Осахаривающая способность еще сильнее выражена у Rhizopus oryzae, применяемом в Японии при изготовлении рисовой водки — саке (алкогольного брожения названная форма не вызывает).
Как распространеннейшие легко культивируемые грибы, мукоровые много изучались экспериментально. Выше уже было указано назначение их в разработке учения о гетероталлизме (стр. Гетероталлизм). Кроме того, они нередко служат объектами для физиологического изучения и особенно для изучения изменчивости и влияния среды на ход развития. В последнем отношении особенно излюбленным объектом служит Phycomyces blakesleeanus.
Contents
Порядок Mucorales может быть разделен на следующие семейства:
- Семейство Мисоrасеае. Органы бесполого размножения представлены типичными многоспоровыми спорангиями почти всегда с колонкой.
- Семейство Chaetocladiaceae. Типичные спорангии постепенно редуцируются и уменьшаются в размерах, превращаясь каждый или в спорангиоль с небольшим числом спор и без колонки, или еще дальше — в односпоровый спорангий или конидию. Наряду с этим у некоторых представителей сохраняются еще и первоначальные многоспоровые спорангии с колонкой (например у Thamnidium).
- Семейство Сhoanephoraceae. Аналогичное предыдущему семейству уменьшение спорангиев с превращением их в спорангиоли или конидии происходит иным путем: вынесением спорообразования на поверхность зачатка спорангия. Наряду с этим у некоторых представителей сохраняются еще и первоначальные многоспоровые спорангии с колонкой (например у Choanephora).
- Семейство Piptocephalidaceae. Такое же, как в предыдущем семействе, вынесение спорообразования на поверхность зачатка первоначального спорангия, но здесь образуются цилиндрические частичные спорангии со спорами, расположенными в один ряд. Они при созревании разламываются на односпоровые членики, так что каждый частичный спорангий биологически превращается в цепочку конидий. Первоначальные многоспоровые спорангии не развиваются.
- Семейство Mortierellaceae. Имеются многоспоровые спорангии, но без колонки. У некоторых вместе с ними или вместо них образуются 1—2 споровые cпорангиоли, или конидии. Зигота окружена обверткой из гиф, образуя зачаточное плодовое тело.
- Семейство Endogonaceae. Как спорангии (без колонок), так и зиготы образуются по нескольку в особых плодовых телах.
1. Семейство Mucoraceae
Главные представители: Mucor, самый распространенный и обширный род среди мукоровых грибов (около 150 видов). Спорангиеносцы простые или ветвящиеся (рис. 125). Спорангии шаровидные. Зиготы голые (без обрастаний со стороны суспензоров). М. mucedo и М. albo-ater — наиболее распространенные виды на навозе и как плесень. К Mucor близок небольшой гомоталличный род Zygorhynchus, отличающийся некоторой гетерогамией; обычен в почве.

1 — Моnоmucor (M. mucedo);
2 — Racemo-muсоr (М. racemosus);
3 — Cymo-muсоr (М. circinelloides);
4 — Sporodinia grandis;
5 — Circinella umbellata;
6 — Thamnidium elegans.
Rhizopus. Характеризуется столонами; на них на месте соприкосновения с субстратом образуются в одну сторону пучки ризоидов, а в другую — группы спорангиеносцев (рис. 22, 2). Спорангии шаровидные. Rh. nigricans — обычная плесень.
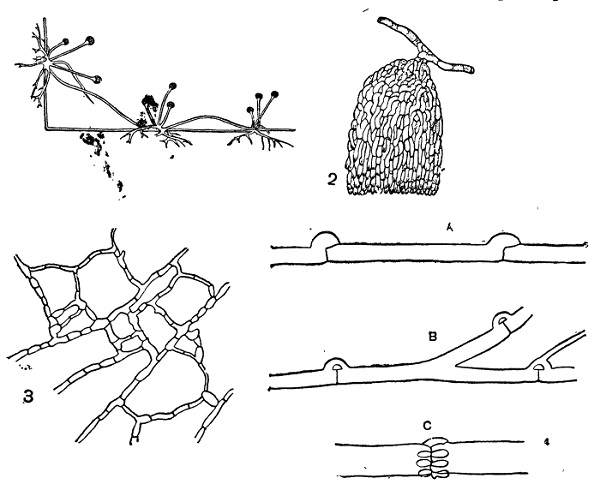
1 — столоны со спорангиеносцами на них и ризоидами;
2 — Sclerotinia tuberosa, органы прикрепления мицелия;
3 — анастомозы мицелия;
4 — пряжки:
А — развитие пряжки;
В — пряжки Merulius;
С — Coniophora.
Absidia. Имеет такие же столоны, как и Rhizopus, но спорангиеносцы образуются по середине дуги, образуемой столонами. Спорангии грушевидные. У большинства на суспензорах образуются выросты, прикрывающие зиготу (рис. 122, 7). У некоторых видов намечается гетерогамия. A. (Thiegemella) glauca обычна в лесной почве.
Phycomyces. Спорангия и спорангиеносцы такого же строения, как у Mucor. На суспензорах развиваются шиловидные отростки, окружающие зиготу. Сюда относятся наиболее крупные среди мукоровых грибов (спорангиеносцы достигают 30 см высоты). Ph. blakesleeanus, встречающийся изредка
на конском навозе, служит одним из излюбленных объектов для экспериментально-генетических исследований (см. стр. Цитология и наследственность).
Sporodinia. Спорангиеносцы, дихотомически ветвящиеся (рис. 125, 4). S. grandis часто встречается как полупаразит на плодовых телах базидиомицетов. Как гомоталличная форма легко дает зиготы.
Pilobolus. Спорангиеносцы не ветвящиеся, вздутые в верхней части, а в нижней обыкновенно отделенные перегородкой от гиф мицелия. В них развивается значительное тургорное давление, которое разрывает оболочку спорангиеносца кольцевым разрывом под колонкой и отбрасывает вместе с ней спорангии на значительное расстояние (рис. 60). P. crystallinus обычен на конском навозе (название crystallinus вызвано блестящими капельками жидкости, выступающей на поверхности спорангиеносца от тургорного давления внутри его).
2. Семейство Chaetocladiaceae.
Thamnidium. Развиваются конечные крупные спорангии с колонкой и мелкие спорангиоли на боковых мутовчато расположенных ветвях спорангиеносца (рис. 126, 1).Th. elegans обычен на навозе и как плесень.
Ghaetocladium. Спороносные ветви многократно ветвятся и на концах образуют конидии (односпоровые спорангиоли). Спорангиев не образуется. Паразиты на других Mucoraceae. Ch. Brefeldii (рис. 126, 2).
3. Семейство Choanephoraceae.
Blakeslea (единственный известный вид — Bl. trispora) образует типичные многоспоровые спорангии с колонкой, а кроме того, — спорангиоли, содержащие обычно только по три споры (рис. 127). Обнаружена в Северной Америке; у нас не найдена.
Choanephora образует многоспоровые спорангии с колонкой и конидии (рис. 127). Последние собраны в головки на конечных вздутиях спороносцев, соответствующих зачаточным спорангиям. Несколько тропических видов; у нас не обнаружена.
Cunninghamella образует только конидии на поверхности шаровидно вздутых конидиеносцев (спороношение типа Oedocephalum (рис. 127, 2)). Большею частью тропические и субтропические; С. albida в почве около Сухуми.
4. Семейство Piptocephalidaceae.
Synceplialastrum. Спороносцы разветвленные, вздутые на конце. На их поверхности образуются головкой цилиндрические спорангии со спорами, расположенными в один ряд. Споры большей частью срастаются боками с оболочкой спорангия (рис. 128, 2).
Syricephalis. Спороносец не разветвленный, вздутый на конце в головку. На ней сидят цилиндрические спорангии со спорами, расположенными в один ряд (рис. 128, 1). Они срастаются с оболочкой спорангия, распадающегося при созревании на отдельные членики. Паразиты на других мукоровых S. sphaerica и др.
Piptocephalis. Спороносцы разветвленные, не вздутые на конце. На них пучками сидят цилиндрические спорангии такого же характера, как у Syncephalis (рис. 128, 3). Паразиты на мукоровых P. fusispora, P. freseniana и др.
5. Семейство Mortierellaceae.
Зиготы оплетены толстым слоем нитей, берущих начало от суспензоров и окружающих вегетативных гиф (зачаточное плодовое тело). Большей частью сапрофиты, иногда полупаразиты на других грибах (особенно мукоровых).
Mortierella образует многоспоровые спорангии без колонки на концах большей частью разветвленных, прямо стоящих спорангиеносцев, оплетенных у основания пучками ризоидов М. candelabrum (рис. 129). Кроме спорангиев, развиваются иногда одиночные конидии с толстой оболочкой (стилоспоры) М. capitata, M. Rostafinskii и др.

Haplosporangium развивает только спорангиоли одиночно на концах разветвленных спороносцев. Они большей частью односпоровые (конидии), но у Н. bisporale — двуспоровые, чем обнаруживают свою природу как спорангиолей (рис. 130).

1 — часть спорангиеносца,
2 — спорангиоль.
6. Семейство Endogonaceae.
Главный и почти единственный род — Endogone с десятком приблизительно описанных видов. Это почвенные грибы с неклеточным мицелием. Довольно вероятно, что в естественных условиях они входят в состав микоризы, специально такой, которая образована гифами без перегородок (см. стр. Микориза). В культуре они не получены. Органы размножения образуются в особых плодовых телах, представляющих большею частью довольно неправильные шарики до 2 см и более в диаметре. Большею частью они развиваются подземно. Известны два сорта плодовых тел: одни с органами бесполого размножения (спорангиями) и другие — с половыми органами и зиготами. Генетической связи между ними не установлено: и те и другие описываются как отдельные виды, однако, весьма вероятно, что в ряде случаев мы имеем здесь только стадии развития одного гриба.
Бесполые плодовые тела представляют довольно плотное сплетение, внутри которого на особых, более толстых гифах как конечные округлые вздутия их образуются спорангии. Они отделяются у основания перегородкой, идущей прямо поперек без образования колонки (как у Моrtierella), и затем развивают споры (рис. 131, 1). Половые плодовые тела такого же в общем строения, как и бесполые. В них развиваются половые органы в виде гиф, расположенных парами параллельно друг другу, с большим количеством клеточных ядер. Дальнейшее развитие их прослежено с достаточной полнотой только у одного вида, Е. lacbiflua. Оно состоит в следующем. Поперечными перегородками на концах указанных гиф отделяется по одной клетке, с одним крупным ядром каждая. Обыкновенно одна из них несколько крупнее другой. Они плотно прикладываются друг к другу, и затем через частичное растворение оболочек содержимое одной клетки (меньшей) переходит в другую (большую). Затем на верхнем свободном конце этой последней появляется вырост, куда переходит ее содержимое вместе с обоими ядрами. Этот вырост и развивается в зигоспору (рис. 131, 2). Его оболочка утолщается, окружающие гифы также характерным образом утолщают свои стенки, в результате чего зигоспора оказывается внутри плодового тела окруженной особой обверткой, напоминающей очертанием огненные языки (рис. 131, 2, с). В таком виде зигоспора переходит в состояние покоя. В это время ядра в ней еще не прокопулировали. Прорастание зиготы неизвестно. Вероятно в этот момент происходит и слияние в ней ядер. У некоторых Endogonе отмечено в плодовых телах образование азигот с такими же «пламенными» обвертками.

1 — спорангий;
2 — образование зигот a, b, c.
Endogone филогенетически довольно близка к Mortierella, представляя дальнейший член того же ряда эволюции. Зачаточные плодовые тела Mortierella, содержащие одну только зиготу, эволюционировали здесь в более массивные образования с большим количеством зигот. Далее, у некоторых Мuсоrасеае наблюдаются сокращение числа ядер в копулирующих гаметангиях (до двух у Zygorrhynchus Dangeardii); здесь у Endogone каждый гаметангий содержит только по одному ядру, т. е. его содержимое соответствует одной только гамете. Что касается бесполых плодовых тел Endogone, то и в этом отношении можно заметить их зачатки у Mortierella в виде густого пучка ризоидов, оплетающих основание спорангиеносца, о чем было упомянуто выше. Однако у Mortierella спорангиеносцы развиваются изолированно, каждый в своей обвертке, тогда как у Endogone они соединяются по нескольку в более массивные плодовые тела.