Contents
Общая характеристика грибов.
Грибы представляют собой очень обширную группу (тип) растений, содержащую до 80 000 описанных видов. Возможно, впрочем, что некоторое количество из указанного числа представляет одни и те же виды, но повторно описанные под разными названиями. Однако, с другой стороны, нужно признать, что видовой состав грибов изучен еще далеко не полно. Особенно это относится к тропическим областям, где, вероятно, будет найдено еще огромное количество новых видов. Поэтому некоторыми микологами-систематиками и флористами высказывается мнение, что грибы по общему числу видов, существующих сейчас на земле, мало чем уступают, а может быть, даже превосходят цветковые растения (около 150 000 видов). При указанном богатстве видами грибы отличаются вместе с тем большим разнообразием их. Стоит только вспомнить, что сюда относятся столь различные представители, как всем известные шляпные почвенные грибы, или древесные трутовики с их крупными сложно построенными плодовыми телами, далее — крупные, иногда огромные дождевики (до 1,5 м в диаметре у американской Calvatia) затем различные подземные клубневидные грибы, как трюфели и др. С другой стороны, к грибам относится еще значительно большее число микроскопических форм, заметных невооруженному глазу в виде различных налетов и плесеней на тех или других субстратах, а нередко и совсем неразличимых без микроскопа.
При всем этом разнообразии можно указать ряд черт, свойственных всем или огромному большинству грибов. Во-первых, все без исключения грибы не содержат хлорофилла и поэтому имеют гетеротрофное питание, т. е. черпают свой углерод только из готовых органических соединений. В соответствии с этим стоит строение их вегетативного тела. За исключением небольшого числа наиболее примитивных видов, живущих как внутриклетные паразиты зеленых растений, почти у всех остальных оно развито в виде мицелия, или грибницы, и представляет систему очень тонких ветвящихся нитей, или гиф, непрерывно нарастающих своими концами и пронизывающих субстрат, на котором развивается гриб. Благодаря такому строению мицелия, он имеет очень большую поверхность, через которую осмотическим путем и поступают в него питательные вещества, в том числе и сравнительно мало подвижные, но необходимые для него органические вещества.
При микроскопическом изучении обнаруживается, что мицелий различно построен у низших и высших грибов. У первых слагающие его гифы, хотя и достигают многих сантиметров в длину, но лишены поперечных перегородок, и, таким образом, весь мицелий оказывается, формально говоря, одной сильно разветвленной клеткой с огромным числом ядер в ней. Такой мицелий называется одноклеточным, или не членистым. У всех высших грибов и немногих низших гифы мицелия перегорожены поперечными перегородками на отдельные клетки, каждая с одним или чаще с несколькими ядрами. Такой мицелий называется многоклеточным, или членистым. За этим, хотя и существенным различием, мицелий всех грибов построен весьма сходно и одинаково пронизывает субстрат или (при большой влажности атмосферы) отчасти растет сверх него, образуя пушистый налет или более объемистые ватообразные скопления большею частью беловатого цвета.
Мицелий построен в общем слишком однообразно, чтобы по нему можно было идентифицировать тот или другой гриб. Более разнообразны и характерны для того или иного вида или группы видов органы размножения, приносящие споры и называемые поэтому также органами спороношения. При этом здесь различают бесполые и половые спороношения. Первые, подобно вегетативному размножению высших растений, служат для быстрого размножения и повторяют, как правило, полностью особенности произведшего их организма. Вторые имеют в основе половой процесс, сопровождающийся слиянием клеточных ядер и следующим затем редукционным делением их. Поэтому споры полового размножения, подобно семенам высших растений, не являются все вполне тождественными генетически друг с другом и с материнским растением.
Половые споры нередко представляют покоящиеся споры и служат в этом случае не столько для непосредственного размножения, сколько для сохранения вида в течение неблагоприятного периода (зима, засуха).
Среди бесполых спор различают эндогенные и экзогенные. Первые имеются только у низших грибов и образуются в большом числе внутри особых клеток, называемых спорангиями, почему и самые споры называют спорангиоспорами. Экзогенные споры, иначе называемые конидиями, свойственны высшим грибам и только немногим низшим. Они образуются не внутри, а на поверхности образующего их органа, называемого конидиеносцем.
Что касается половых спор, то они также бывают различны. У низших грибов они имеют характер покоящихся спор и различаются по форме полового процесса, давшего им начало (ооспоры, зигоспоры). При прорастании их развивается спорангий со спорами в нем, происшедшими в результате редукционного деления (т. е. генетически не однородными). У высших грибов половой цикл сложнее. Продукт оплодотворения у них не превращается в покоящуюся спору, а разрастается далее и в конце своего развития приносит многочисленные спороносные органы. Одни из них образуют внутри эндогенные споры и называются сумками, или асками, а их споры — аскоспорами. Другие образуют на своей поверхности экзогенные споры и называются базидиями, а их споры — базидиоспорами. Как аскоспоры, так и базидиоспоры образуются в результате редукционного деления, т. е. являются генетически неоднородными, как это свойственно вообще спорам полового происхождения (см. ниже споры спорангиев, получающихся при прорастании половых спор низших грибов).
Для пояснения вышеизложенного можно рассмотреть несколько конкретных примеров.
Обычнейший плесеневый гриб, Mucor mucedo, принадлежит к низшим грибам. Его бесполые спорангиоспоры выпадают из спорангия и, перенесенные токами воздуха на субстрат подходящего состава и влажности, немедленно прорастают. При этом спора вздувается и затем из нее вырастают 2—3 ростковые трубочки, которые быстро растут своими концами дальше, ветвятся и развиваются в нечленистый (одноклеточный или, лучше, неклеточный) мицелий (рис. 1). Он отчасти пронизывает субстрат, отчасти стелется по его поверхности. Через 2—3 дня роста на таком мицелии развиваются особые ветви, которые не ветвятся и растут прямо вверх на высоту до 10 см. Они представляют спорангиеносцы. Их концы шаровидно вздуваются, отделяются от спорангиеносца перегородкой и превращаются в спорангии (рис. 1). В спорангиях без всякого редукционного деления образуются спорангиоспоры таким образом, что общее протоплазменное содержимое распадается на большое число участков, одевающихся каждый собственной оболочкой (рис. 1, 2). Спорангиоспоры, выпадая вследствие разрушения оболочки спорангия, немедленно могут прорастать в такой же мицелий с новыми спорангиями на нем и т. д.
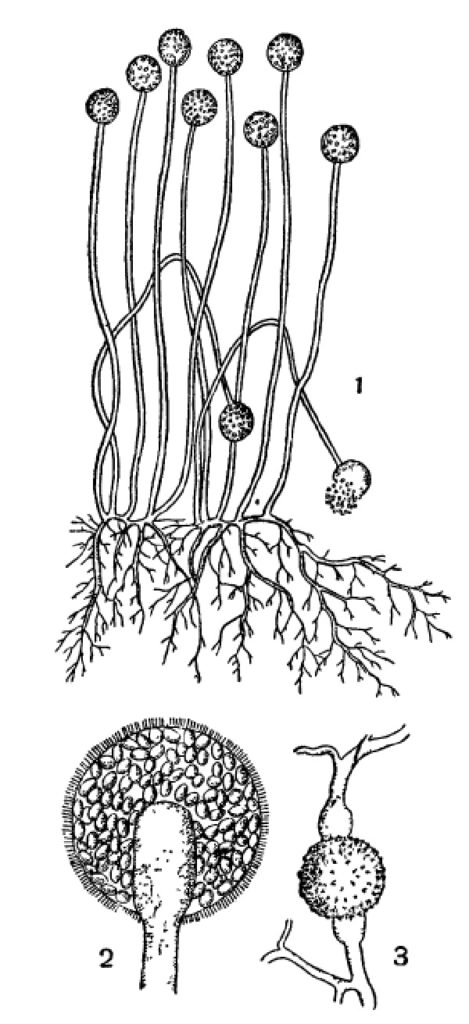
1 — спорангиеносцы со спорангиями и мицелий;
2 — спорангий, сильно увеличенный;
3 — зигоспора на месте слияния двух одинаковых ветвей мицелия.
У Mucor mucedo имеется и половой процесс. Он происходит лишь при определенных условиях: при встрече двух различных в половом отношении мицелиев. При этом специальные короткие ветви гиф того и другого сливаются «своими концами, и из этого слияния получается зигоспора, одевающаяся толстыми оболочками и переходящая в состояние покоя (рис. 1,3). Позднее она прорастает в короткую нить с спорангием на конце. В нем на этот раз происходит редукционное деление при формировании спорангиоспор, так что последние в этом так называемом зародышевом спорангии оказываются генетически разнородными, отличаясь, между прочим, половыми свойствами развивающихся из них мицелиев.
Другой также очень обычный плесеневый гриб, Aspergillus, принадлежит к высшим грибам и имеет членистый многоклеточный мицелий. На нем развиваются конидиеносцы в виде вертикально растущих ветвей длиной в 1— 2 мм, вздувающихся на конце. На поверхности этого вздутия образуются короткие цилиндрические выросты, отчленяющие на своих концах последовательно целые цепочки конидий (рис. 2). Последние отпадают и, попав на подходящий субстрат, легко прорастают, давая 2—3 ростковые трубочки, нарастающие концами и ветвящиеся. В отличие от Mucor, они вскоре делятся поперечными перегородками, так что получается многоклеточный мицелий. На нем опять вскоре формируются такие же конидиеносцы и т. д.

1 — конидиеносец с цепочками конидий;
2 — половые органы;
3 — развитие плодового тела (спиральное тело, в середине — разросшийся женский половой орган с зачатками аскогенных гиф на нем);
4 — часть разреза через аскокарп с сумками и аскоспорами в них.
На том же мицелии развивается и половое спороношение. Вначале появляются половые органы в виде парами расположенных коротких ветвей, часто одной и той же гифы. Одна из этих ветвей, спирально закрученная, является женским органом, а другая, более прямая — мужским (рис. 2, 2). Вскоре затем женский орган дает боковые короткие выросты, называемые аскогеными гифами. На их концах формируются сумки, а в них сначала происходит слияние двух имеющихся ядер и затем редукционное деление копуляциояного ядра. После следующих затем еще двух делений в сумке оказывается восемь ядер, и около них в каждой сумке формируется по восемь аскоспор. Последние, как происходящие в результате редукционного деления, оказываются такими же неоднократными в генетическом отношении, как и споры Mucor в зародышевом спорангии.
Одновременно с указанным развитием половых органов происходит обрастание их соседними ветвями мицелия, которые плотно сплетаются в округлое тельце, и уже внутри него развиваются аскогенные гифы и сумки. Аскоспоры освобождаются вследствие разрушения оболочки этого тельца, называемого сумчатым плодовым телом, или аскокарпом (рис. 2, 4).
Третим и последним примером можно взять какой-нибудь из шляпных грибов, например часто развивающийся на навозе Coprinus (рис. 3). На его многоклетном мицелии могут развиваться, хотя и не особенно обильно, бесполые (конидиальные) спороношения, а затем залагаются плодовые тела, или базидиокарпы, состоящие из пенька и шляпки. На нижней стороне последней развиваются в очень большом числе спороносные органы — базидии. Так же как и сумки, они имеют сначала по два ядра, которые затем сливаются, и после этого копуляционное ядро делится редукционным путем. Затем следует еще одно деление, и в базидии получается четыре ядра. Тем временем на ее поверхности образуются четыре выроста, и в них переходит по одному ядру (рис. 3, 2). Таким образом, на каждой базидии образуется по четыре экзогенных споры, называемых базидиоспорами. По характеру своего развития, при котором происходит слияние клеточных ядер и затем редукционное деление, они сходны с аскоспорами и должны быть отнесены к половым спороношениям. Как те, так и другие, в отличие от бесполых спор, оказываются генетически неоднородными.

1 — плодовое тело с остатками диплоидного мицелия у основания ножки;
2 — базидии (b) с 4 базидиоспорами каждая.
Полиморфизм.
Из всех указанных органов — мицелия и органов спороношения — первый построен настолько однообразно, что идентификация гриба по нему большей частью совершенно невозможна. Характерные признаки приходится поэтому искать во вторых. Однако один и тот же гриб может иметь их несколько: половые спороношения и бесполые, притом последних нередко целый ряд (до 5—6) и иногда весьма различных. Благодаря этому гриб может в корне менять свою физиономию на разных стадиях, так что, если бы не знать генетической связи между ними, то их пришлось бы принять за совершенно разные организмы. Раньше обыкновенно так и делалось. Например, конидиальные спороношения разобранного выше в качестве примера Aspergillus назывались Aspergillus glaucus, а сумчатые — Eurotium herbariorum. Также для представителей современного рода ржавчинных грибов, Puccinia, одно спороношение носило название Spermogonium, другое — Aecidium, третье — Uredo, четвертое — собственно Puccinia, — и все это не в качестве стадий развития одного и того же организма, а в качестве самостоятельных даже родов. Позднее, при изучении полного цикла развития и установлении генетической связи спороношений, эти названия утратили прежний смысл и, если сохранились иногда, то лишь для обозначения того или иного спороношения (например, в приведенном выше примере ржавчинных грибов — названия Aecidium, Uredo и др.).
Это явление — способность грибов менять свой внешний вид в состоянии разных спороношений — получило название полиморфизма, или плеоморфизма их.
Значение для идентификации грибов разных спороношений.
Ввиду указанного полиморфизма встает вопрос, насколько то или иное спороношение может характеризовать данный гриб. Здесь приходится различать органы бесполого и полового спороношений. Поскольку дело касается первых (бесполых), то в некоторых случаях они вполне типичны для данного круга форм и не повторяются у других. Таковы, например, спорангии у мукоровых.
То же, пожалуй, хотя и не с такой точностью, можно сказать относительно конидиеносцев Aspergillus и некоторых других. Однако очень часто бесполые спороношения не достаточно характерны для установления систематического положения гриба. Таковы, например, конидиеносцы так называемого типа Oedocephalum, похожие отчасти на Aspergillus, но отличающиеся тем, что конидии сидят на вздутом кончике конидиеносца не цепочками, а в один только этаж. Они встречаются у Peziza vesiculosa из аскомицетов, у Fomes annosus из базидиомицетов (трутовиков), у Cunninghamella из мукоровых, у Basidiophora из пероноспоровых, а, кроме того, существует еще немало форм с такими же конидиеносцами, у которых неизвестны, кроме них, иные спороношения и которые причисляются поэтому к группе несовершенных грибов (Fungi imperfecti) (рис. 4, 2). Другой пример: похожие на Oedocephalum, но не вздутые на конце конидиеносцы типа Botrytis известны у многих несовершенных грибов (не имеющих, кроме них, других спороношений); кроме того, они развиваются как бесполое спороношение у сумчатого гриба Sclerotinia fuckeliana (рис. 4, 2), тогда как у многих других видов того же рода Sclerotinia бесполые спороношения совсем иного рода, в виде так называемой Моnilia (цепочка конидий на коротком конидиеносце). Если бы основываться здесь на бесполых спороношениях, то пришлось бы разбить очень характерный и хорошо отграниченый род Sclerotinia. Наоборот, половые спороношения оказываются очень характерными не только для идентификации данного гриба, но и для установления его филогенетических отношений. Поэтому они кладутся в основу изучения, и на основании их производятся систематические
группировки.

1 — типа Oedocephalum;
2 — типа Botrytis.
Половое воспроизведение.
Ввиду того, что половое воспроизведение грибов весьма своеобразно и у многих изменило свою первоначальную форму, для понимания необходимо вкратце остановиться здесь на его цитологической основе. Типичный половой процесс в основе своей имеет слияние двух клеток (гамет) и их клеточных ядер. Получившееся при этом копуляционное ядро содержит двойное число хромосом и называется диплоидным. В известный момент развития, разный для разных организмов, диплоидное ядро претерпевает редукционное деление, которым снова восстанавливается гаплоидное число хромосом. Таким образом, для каждого организма, имеющего половой процесс, характерно, что в его цикле развития имеется чередование гаплоидного и диплоидного состояний, или ядерных фаз (от копуляции до редукции — диплоидная фаза, от редукции до новой копуляции — гаплоидная).
Если подойти с этой точки зрения к половому воспроизведению грибов, то можно заметить следующее: у низших грибов (архимицетов и фикомицитов) непосредственным результатом оплодотворения является зигота, имеющая характер покоящейся споры. При прорастании ее копуляционное диплоидное ядро (или ядра, если как у Mucor оплодотворение заключалось в слиянии двух многоядерных клеток) делится редукционным способом, благодаря чему восстанавливается гаплоидное число хромозом. Таким образом, здесь диплоидная фаза представлена только покоящейся зиготой, и вся активная жизнь гриба проходит в гаплоидном состоянии.
У аскомицетов мы также наблюдаем оплодотворение. Однако при этом за слиянием половых клеток, или цитогамией (вернее плазмогамией) не следует сейчас же слияния их ядер, или кариогамии. Мужские и женские ядра в оплодотворенном женском половом органе (архикарпе) только сближаются попарно друг с другом, образуя так называемые синкарионы, или дикарионы.
При разрастании половых органов и развитии из них аскогенных гиф дикарионы размножаются, причем в ряде случаев прослежено, что оба ядра делятся строго одновременно и параллельно друг другу. В молодой сумке происходит слияние этих двух ядер дикариона в одно копуляционное диплоидное ядро. Оно затем сейчас же претерпевает редукционное деление, так что аскоспоры содержат уже гаплоидные ядра и прорастают в гаплоидный мицелий. Часть этого цикла развития от плазмогамии до кариогамии может быть отождествлена с диплоидной фазой, так как каждый содержащийся здесь дикарион ведет себя как нечто единое и содержит двойное число хромосом. Поэтому по существу является второстепенным моментом, что они заключены здесь в две ядерные оболочки, а не в одну. Таким образом, у аскомицетов чередование ядерных фаз выражается в следующем: от аскоспор до плазмогамии идет гаплоидная фаза; от плазмогамии до кариогамии, в сумке — диплоидная фаза, или, как ее называют еще по характерному строению ядерного аппарата, дикариофит. В отличие от низших грибов диплоидная фаза представлена здесь не покоящимся состоянием (зигоспорой), а имеет известное разрастание и размножение своих ядер. Однако преобладание в цикле развития имеет все-таки гаплоидная фаза, так как на ее долю приходится не только вегетативный мицелий, могущий неограниченно размножаться путем образования бесполых спороношений, но и значительная часть сумчатого плодового тела (вся оболочка его и часть внутренней массы, кроме аскогенных гиф и молодых сумок). Нужно отметить еще, что у многих аскомицетов слияния половых клеток совсем не происходит, но всегда тем или иным образом (апогамия) происходит образование дикарионов и развитие аскогенных гиф, так что характер смены ядерных фаз при этом по существу не изменяется.
Что касается, наконец, базидиомицетов, то у них совершенно неизвестно образования половых клеток. Базидиоспоры здесь, как и аскоспоры, гаплоидны. При прорастании они дают гаплоидный, или первичный мицелий, который может размножаться бесполым путем, но в общем имеет непродолжительное существование и вскоре заменяется диплоидным, или вторичным мицелием. Последний возникает в результате анастомозов и слияний гиф одного первичного мицелия или чаще двух разных. Через анастомоз переходит ядро из одной клетки в другую, в результате чего получаются первые двуядерные клетки с дикарионами. Они затем разрастаются в целый мицелий, сложенный из таких двуядерных клеток (при этом, конечно, дикарионы размножаются подобно тому, как и в аскогенных гифах). Этот вторичный мицелий, относящийся к диплоидной фазе (дикариофит), обычно имеет более продолжительное существование и часто бывает даже многолетним. За счет его полностью слагается все плодовое тело. В развива юпщхся на нем базидиях, так же как и в сумках, происходит, наконец, слияние ядер дикариона, и копуляционное ядро сейчас же делится затем редукционным образом, так что развивающиеся на базидии базидиоспоры содержат уже по одному гаплоидному ядру. Базидиоспора снова прорастает в первичный гаплоидный мицелий. Таким образом, и у базидиомицетов мы имеем ясную смену ядерных фаз: от базидиоспоры до возникновения первых дикарионов — гаплоидная фаза; от первых дикарионов через весь вторичный мицеЛий и все плодовое тело до базидий — диплоидная фаза, цитологически представленная, как и у аскомицетов, в виде дикариофита, но в отличие от них здесь преобладающая. Из приведенного видно, что базидиальное спороношение, подобно сумчатому стоящее в связи со сменой ядерных фаз (рис 5), является по существу выражением половой функции. Поэтому оно вместе с половыми спороношениями других грибов привлекает особое внимание миколога, так как здесь находят наиболее надежные указания на филогенетические отношения и эволюцию изучаемых групп.

1 —у фикомицетов;
2 — у аскомицетов;
3 — у базидиомицетов. Тонкий контур — гаплоидная фаза, жирный—диплоидная фаза.
Однако было бы односторонним ограничиваться только статическим изучением половых спороношений. Истинная природа всякого организма лучше выясняется в его динамике, особенно в индивидуальном развитии, или онтогенезе. Особенное значение имеет такой подход для грибов с их часто очень сложным циклом развития. Поэтому современное морфолого-систематическое изучение грибов, дающее в первую очередь основу для понимания их истинной природы и филогенетических взаимоотношений, ставит перед собой более сложные задачи: выяснение полного цикла развития с учетом цитологических явлений и сравнительное изучение не только строения, но и развития как вегетативных органов, так и различных спороношений, образующихся в той или другой фазе развития.