Этот порядок наиболее типичен среди базидиомицетов и вместе с тем наиболее обширен (около 12 тысяч видов). Как показывает название, наиболее характерным признаком считается здесь расположение базидии гимением на поверхности плодового тела. Лишь у немногих простейших представителей гимений зачаточный или совсем не выражен, так же как и плодовые тела. В типичном случае гимений состоит из слоя цилиндрических или слегка расширенных на вершине (булавовидных) базидии и чередующихся с ними парафиз, обыкновенно такой же формы и также в молодости содержащих два ядра, но не сливающихся затем (рис. 235). Реже и преимущественно у простейших представителей парафизы нитевидные, иногда даже ветвящиеся (у видов Aleurodiscus).
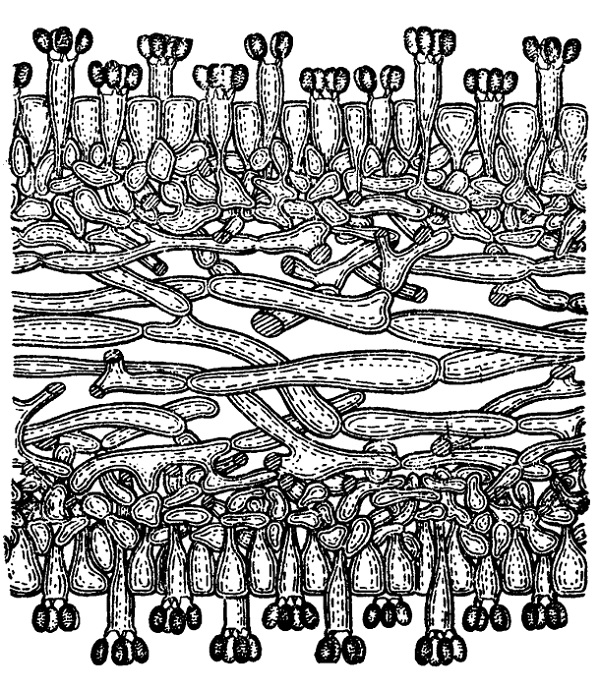
У некоторых гименомицетов развиваются еще особые цистиды в виде более крупных, чем парафизы, образований, возвышающихся своими концами, над общим уровнем гимения. Их стенки нередко утолщены или инкрустированы, и они играют главным образом защитную роль, предохраняя молодой гимений от соприкосновения с другими поверхностями, что могло бы повредить развивающиеся базидиоспоры. Что же касается парафиз, то как сочные тургесцентные образования они придают известную упругость всему гимениальному слою и поддерживают с боков базидии, отделяя их вместе с тем друг от друга и предохраняя от слипания сидящие на них базидиоспоры.
Базидии всегда построены по акроспоровому типу, т. е. несут базидиоспоры на своей вершине (почти всегда по 4). Последние по созревании отбрасываются с некоторой силой от поверхности гимения, причем это происходит большей частью не одновременно для всех четырех базидиоспор. После отделения спор базидия отмирает и спадается. Согласно прежним указаниям (Juel, 1898, и др.), там, где как у Cantharellus в базидии в результате трех делений получается 8 ядер и из них 4 переходят в базидиоспоры, а 4 остаются в самой базидии — после отделения первой генерации базидиоспор развивается затем вторая. Однако новейшие исследования (главным образом Buller) не подтверждают этого. Базидия развивает только одну генерацию спор и отмирает вслед за их отделением. У большинства гименомицетов отделение спор с гимения происходит постепенно, так что на одном небольшом участке находятся базидии разного возраста, причем по мере отмирания более старых между ними из более глубоких слоев выдвигаются новые и начинают развивать на своей вершине базидиоспоры. Таким образом, во многих случаях развивается до 4—5 последовательных генераций базидии (например у шампиньонов и многих других Agaricaceae). У некоторых трутовиков гимений функционирует в течение всего вегетационного периода, а у некоторых оказывается даже многолетним (например у Daedalea). Надо думать, что здесь число генераций последовательно образующихся базидии должно быть весьма значительным. Реже встречается другой тип, представленный главным образом у видов Goprinus. Здесь определенный участок гимения функционирует только очень непродолжительное время и после отделения базидиоспор весь отмирает и расплывается. Тем временем тот же процесс переходит на соседний участок и т. д. Однако и у Goprinus с его столь эфемерным гимением удается различить в нем два сорта базидии: более длинные и более короткие. Они развиваются почти одновременно, но все-таки первые раньше сбрасывают свои споры, чем вторые (рис. 235).
Гимений развивается на плодовых телах мясистой, кожистой или деревянистой консистенции. У низших представителей — плодовые тела в виде поверхностного плоского сплетения гиф, неопределенно нарастающего своими краями. Его верхняя свободная поверхность несет гимений, также не ограниченный в своих очертаниях. Такие плодовые тела называются ресупинатными. Они типично представлены, например у Coniophora, Соrticium и некоторых других (из семейства Thelephoraceae). У близкого систематически Stereum такие же плоские плодовые тела обыкновенно имеют приподнимающиеся от субстрата края, и гимений развивается здесь только на их нижней поверхности. Здесь мы видим характерную вообще для гименомицетов ориентировку гимения на нижнюю сторону, что стоит в связи с характером распространения базидиоспор. У относящихся к тому же семейству Thelephora и некоторых других плоские плодовые тела часто стоят почти вертикально, прикрепляясь к субстрату суженным основанием. Гимений здесь — или с обеих сторон или только с одной, обращенной книзу. Наконец, у Crater ellus и некоторых других плодовые тела построены по радиальному типу и имеют вид воронки с узким нижним концом. Гимений находится здесь на наружной, отчасти книзу обращенной поверхности.
Аналогичную эволюцию плодовых тел от ресупинатных плоских до вертикально стоящих и радиальных можно проследить и в других семействах гименомицетов (Hydnaceae, Polyporaceae), причем параллельно с этим изменением формы также наблюдается перенесение гимения на нижнюю сторону.
Другая существенная черта строения плодовых тел гименомицетов заключается в расположении на них гимения. У одних, преимущественно низших форм, несущая гимений поверхность, или гименофор, не выделяется особенно от остальной поверхности плодового тела и оказывается гладкой или слегка бородавчатой (гименофор гладкий). У других гимений располагается на особых выростах плодового тела, имеющих определенную форму (гименофор в виде шипов, трубочек или пластинок). Эти различия кладутся в основу систематического разделения гименомицетов.
Наконец, весьма существенно то, что у одних гимений с самого начала лежит открыто на поверхности плодового тела или его гименофоров, у других же он развивается сначала в замкнутой полости, прикрытый сплетением стерильных гиф, называемых покрывалом (velum), и обнажается лишь при созревании базидий вследствие разрыва или разрушения этого покрывала. Первый тип может быть назван гимнокарпным, а второй гемиангиокарпным. К первому относятся более примитивные группы (Thelephoraceae, Glavariaceae, Hydnaceae и частью Polyporaceae), а второй представлен у высших Polyporaceae (Boletus) и особенно у Agaricaceae. Плодовые тела у этих последних обычно состоят из пенька и шляпки, причем шляпка в молодости прижата свободным краем к пеньку, и здесь между ними существует соединение, образованное покрывалом. Позднее, при развертывании гимения, оно разрывается. У некоторых Agaricaceae, как у Amanita, кроме этого покрывала, называемого частным (velum partiale), имеется еще общее (velum universale), которое в молодости окружает все плодовое тело и разрывается раньше частного покрывала вследствие вытягивания пенька. При этом часть его остается в виде оторочки у основания пенька, а другая часть остается на шляпке в виде лоскутов (такое именно происхождение имеют белые пятна на верхней поверхности шляпки обычного красного мухомора).
Анатомическое строение плодовых тел
При нередко значительных размерах и сложности внешнего расчленения плодовые тела гименомицетов могут достигать также и довольно значительной сложности анатомического строения. Кроющая ткань диференцирована преимущественно на верхней стороне шляпки у Agaricaceae и некоторых Polyporaceae. О строении кроющей ткани см. стр. Кроющая ткань. Нередко она может быть отодрана в виде более темно окрашенной пленки от более светлой и рыхлой внутренней части. Последняя представляет или войлочное сплетение гриф, или в некоторых мясистых плодовых телах носит параплектенхиматический характер. У некоторых Agaricaceae, как у Lactarius, эта внутренняя ткань состоит из гнездами расположенных параплектенхиматических участков и отделяющих их друг от друга гифенных войлочных прослоек. Весьма широко распространены в плодовых телах элементы, проводящие органические вещества (см. рис. 19); у Lactarius и некоторых других они имеют характер млечных трубок, у большинства же просто отличаются более густым содержимым, богатым жиром и другими питательными веществами. Эти проводящие гифы особенно обильно подходят к гимению и иногда высовываются своими концами на поверхность, образуя своеобразные как бы цистиды (глеоцистиды) (рис. 18, 1,2). У трутовиков, как виды Fomes,Trametes и др., плодовые тела отличаются значительной твердостью. Это обусловлено тем, что слагающие их гифы обладают сильно утолщенными прочными стенками, но обыкновенно не замечается ясной группировки этих имеющих механическое значение гиф в определенные механические ткани. Лишь кое-где в пеньках шляпных грибов замечается скопление утолщенных элементов на периферии (кроющая ткань в пеньках большею частью не выражена с достаточной ясностью). Чаще всего упругость и прочность пенька на изгиб обусловлена преимущественно тургором слагающих его гиф и известным напряжением, какое существует между сжатой в продольном направлении внутренней его частью и растянутой наружной. Определенной эмбриональной ткани в плодовых телах гименомицетов не имеется, если не считать за таковую краевой нарастающей зоны у ресупинатных форм. Обычно рост плодового тела обусловлен более или менее равномерным разрастанием всех его частей, хотя иногда отмечается более сильный рост в верхних участках пенька у шляпных грибов (интеркалярная зона роста).
Плодовые тела мясистой консистенции обыкновенно однолетние и часто существуют в развитом состоянии всего несколько дней; что же касается кожистых и особенно деревянистых плодовых тел, то они обычно многолетние. При этом у одних, как Schizophyllum, Daedalea, Trametes и др., все время сохраняется один и тот же гимений, прерывающий только свою жизнедеятельность на время зимы или сухого периода. У других, как представители рода Fomes, гимений функционирует только один сезон, а на следующий год поверх него развивается новый и т. д. Такие плодовые тела, имеющие ясно выраженное зональное нарастание, не только с нижней гимениальной стороны, но и сверху, достигают иногда очень значительных размеров и солидного возраста (до 80 лет).
По образу жизни большинство гименомицетов относится к почвенным макрофитам. Значительное количество их также живет в качестве сапрофитов на древесине или паразитов на стволах деревьев. Общая биология тех и других освещена в 1-й части (см. стр. Древесные грибы, Копрофильные грибы, Почвенные грибы), так же, как и распространение у них спор и соответственно с этим строение и расположение гимения и гименофоров (см. стр. Распространение спор).
Семейства гименомицетов
Порядок Hymenomycetales может быть разделен на следующие семейства:
- Гимений расположен непосредственно на поверхности плодового тела (гименофор гладкий).
а) Плодовые тела зачаточные, паутинистые или у большинства ясно развитые, кожистой консистенции, большей частью плоские, иногда прямо стоящие, воронковидные. Гимений с одной стороны. - Сем. Thelephoraceae.
б) Плодовые тела мясистые, прямо стоящие, булавовидные или ветвящиеся. Гимений большей частью покрывает их со всех сторон. - Сем. Clavariaceae.
- Гимений расположен на особых выростах плодового тела (гименофорах).
а) Гименофор в виде шипов или зубцов. - Сем. Hydnaceae.
б) Гименофор в виде трубок или переплетающихся складок. - Сем. Polyроrасеае.
в) Гименофор в виде пластинок, расходящихся радиально от центра (большей частью от пенька). - Сем. Agaricaceae.
Приводимое здесь разделение при всей его внешней простоте и наглядности практически может иногда создать затруднение, так как существуют формы, обнаруживающие промежуточный характер между указанными семействами; поэтому провести резкое разграничение между ними не всегда возможно. С другой стороны, эта система оспаривается некоторыми с точки зрения филогении. Здесь некоторые современные авторы придают преимущественное значение, с одной стороны, строению и развитию базидии (стихо- и хиастобази-диальные), а с другой — гимнокарпности и гемиангиокарпности. Относительно значения первого момента некоторые соображения были высказаны выше (см. стр. Базидиомицеты). Здесь можно только указать, что история развития базидий прослежена у такого сравнительно ничтожного количества форм, что полагать ее в основу систематического разделения всей огромной группы гименомицетов кажется слишком смелым, тем более, что у близких по остальным свойствам видов встречается как тот, так и другой тип развития (например у разных видов Clavaria). Что касается второго, то при всей принципиальной важности его мы также располагаем еще слишком скудными данными относительно истории развития плодовых тел и заложения гимения, чтобы класть этот принцип в основу систематики. Сторонники его, Maire и за ним Гейман, объединяют, например, в свои Agaricales огромное количество видов, которые они считают гемиангиокарпными. При этом сюда относятся не только пластинчатые грибы, но и Boletus. Все остальные считаются гимнокарпными. Однако фактически для огромного большинства тех и других мы совершенно не знаем этой стороны развития. Во всяком случае принятая здесь система имеет преимущество простоты и наглядности, хотя и нельзя отрицать, что, может быть, она не совсем точно иногда отражает филогенетические отношения.