У архимицетов для некоторых форм установлена копуляция одинаковых планогамет (подвижных гамет), не отличающихся по существу от зооспор бесполого размножения (некоторые виды Olpidium, Synehytrium endobioticum; рис. 33, 1). В других случаях наблюдается гологамия, т, е. слияние в половом процессе двух вегетативных особей, не диференцированных как специальные гаметы. При этом они или непосредственно прикладываются друг к другу, и содержимое одной (часто меньшей) переливается в другую (Sporophlyctis, некоторые Chytridium), или между ними образуется более или менее длинный копуляционный отросток (Zygorhyzidium, Polyphagia.) Соответственно общему строению Mycochytridiales сливающиеся особи здесь одноядерны, но у Olpidiopsis (из Myxochytridiales), где имеется такое же слияние, они многоядерны и могут быть сравнены с целыми гаметангиями, в которых только не произошло обособления отдельных гамет.
Среди оомицетов наиболее примитивным в смысле полового процесса является Allomyces javanicus, недавно изученный Книпом (1929). Здесь наблюдается анизогамия — слияние двух подвижных гамет неодинаковой величины (♂ и ♀). По строению они (особенно ♀) сходны с зооспорами бесполого размножения, но отличаются от них физиологически (рис. 33, 2). Allomyces и, может быть, другие Blastocladiaceae, которые, кроме названного примера, не изучены в половом отношении, представляют дальнейший этап эволюции изогамных хитридиевых (как Olpidium). У остальных оомицетов имеется резко выраженная оогамия. У Monoblepharis женская гамета — яйцо в виде одноядерной шаровидной клетки свободно лежит в оогонии и даже иногда высовывается из него. Мужские гаметы представлены сперматозоидами — подвижными одножгутиковыми клетками, образующимися по несколько штук в антеридии. Оплодотворение состоит в слиянии сперматозоида с яйцом (рис, 33, 3). У остальных оомицетов (и вообще у всех остальных грибов) обособленных сперматозоидов не образуется; мужской орган представлен здесь антеридием, почти всегда многоядерным и поэтому сравнимым с целым гаметангием, но без диференцировки на отдельные гаметы. Женский орган — оогоний — построен не везде одинаково. У Saprolegniaceae он закладывается как многоядерная клетка, поэтому также сравним с гаметангием. Затем в нем происходит отмирание значительного числа ядер, и протоплазма распадается на некоторое число одноядерных яйцеклеток. Оплодотворение состоит в том, что антеридий пускает отростки, прободающие оболочку рогония (оплодотворяющие отростки); они ветвятся и концами прикладываются к яйцеклеткам, переливая затем в них часть своего содержимого с одним ядром; позднее оно сливается с ядром яйцеклетки (рис. 33, 4). У Peronosporales и некоторых Saprolegniales, например Araiospora, в оогоний диференцируется всегда только одно яйцо, на образование которого идет, однако, не вся протоплазма, а лишь центральная часть ее, периферическая же часть остается в виде так называемой периплазмы и служит для питания яйцеклетки и образования затем толстой оболочки около ооспоры. Оогоний здесь также залагается как многоядерная клетка и может быть сравнен с гаметангием. При диференцировке в нем яйцеклетки эта последняя в большинстве случаев получает только одно ядро, а остальные отходят в периплазму (рис. 33, 5); однако у Albugo bliti и Albugo portulacae яйцеклетка оказывается многоядерной. При оплодотворении у большинства в яйцеклетку переходит только одно мужское ядро, сливающееся с женским, а у Albugo bliti и A. portulacae таким же образом переходит много мужских ядер, которые и сливаются попарно с имеющимися там женскими ядрами.
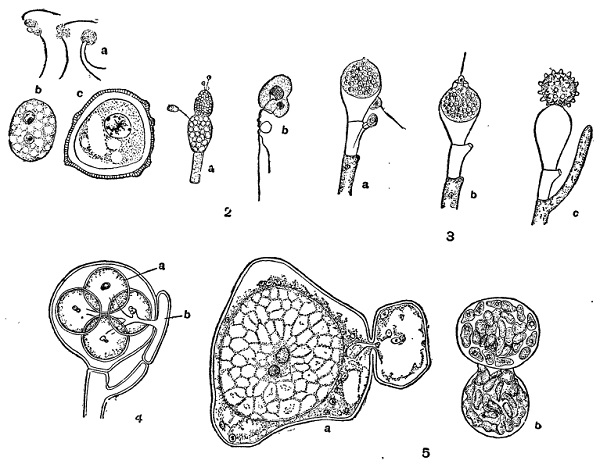
1 — Olpidium viciae:
a — копуляции гамет;
Ь и с — молодая и взрослая зиготы.
2 — Allomyces javanicus:
а — мужской и женский гаметангии;
b — копуляция гамет.
3 — Monoblepharis sphaerica:
а и b — оплодотворение;
с — ооспора.
4 — Saprolegnia:
а — яйцеклетки;
b — антеридий.
б — Albugo Candida:
a — оплодотворение;
b — прорастание ооспоры.
У зигомицетов половой процесс, называемый зигогамией, заключается в слиянии двух большею частью многоядерных клеток — гаметангиев, содержимое которых лишено диференцировки, какая хотя бы имеется в оогониях оомицетов. Они образуются обыкновенно на концах особых коротких ветвей мицелия (копуляционных ветвей) и большей частью совершенно одинаковы по величине и строению (изогамия) (рис. 34, 2). Однако иногда наблюдаются некоторые различия; например у Zygorhynchus или Absidia (из мукоровых) и некоторых других одна клетка всегда больше другой, так что здесь получается некоторый зачаток анизогамии (гетерогамии) (рис. 34, 2). Наконец, у Endogone и Basidiobolus сливающиеся клетки формируются как одноядерные, и в зиготе происходит слияние только одной пары клеточных ядер. Таким образом, и у зигомицетов в эволюции полового процесса происходит замена множественного оплодотворения при слиянии двух гаметангиев простым соединением одноядерных элементов, которые можно сравнить с гаметами, хотя и лишенными диференцировки (рис. 34, 3).
Хотя прорастание половых покоящихся спор у низших грибов (архимицетов, оомицетов и зигомицетов) изучено цитологически очень недостаточно, но все-таки в некоторых случаях установлено, что оплодотворенное диплоидное ядро делится при прорастании редукционным путем (например у Olpidium viciae, — Kusano, 1912; Saprolegnia monoica, — Claussen, 1908; Plasmopara viticola, —Arens, 1929). В других случаях за это говорят генетические данные: разделение полов и расщепление признаков гибридов (у многих мукоровых). Поэтому полагают, что при прорастании споры, происшедшей половым путем, имеет место редукция и возвращение к гаплоидному состоянию.
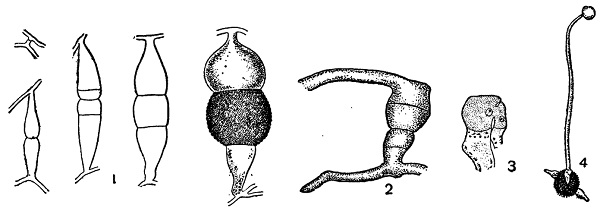
1 — Rhizopus nigricans;
2 — Absidia spinosa — гетерогамная копуляция;
3 — Endogone — гетерогамная копуляция одноядерных клеток;
4 — Мuсоr—прорастание зигоспоры образованием зародышевого спорангия.
Необходимо отметить, наконец, что покоящиеся споры, сходные с половыми, образуются иногда и без всякого оплодотворения. Такое явление имеет место и среди хитридиевых и особенно известно среди мукоровых и сапролегниевых. У мукоровых иногда развиваются так называемые азиготы — или парой, на месте подошедших друг к другу, но не слившихся гаметангиев, или одиночно, на отдельно стоящей копуляционной ветви. У многих сапролегниевых антеридии образуются далеко непостоянно, а у некоторых видов совершенно отсутствуют; тем не менее в их оогониях яйцеклетки развиваются партеногенетически как бы в настоящие ооспоры (партеноспоры). Возникающие таким путем споры прорастают также после периода покоя совершенно сходным образом с половыми спорами, но, как нужно думать, без редукционного деления ядер.