К этому семейству относится более 500 видов, главным образом сапрофитов на древесных остатках, иногда на почве.
У простейших представителей плодовых тел по существу совсем не образуется, и базидии располагаются на очень рыхлом паутинистом сплетении гиф, не смыкаясь в гимений; иногда даже совсем одиночно. Таковы представители рода Hypochnus (рис. 236). От Hypochnus имеются переходы к Corticium. У большинства представителей этого обширного рода имеется плодовое тело ресупинатного типа в виде войлочного более или менее плотного сплетения гиф на поверхности субстрата, несущего на верхней стороне гимений (образованный только базидиями без парафиз). Однако у простейших Gorticium это плодовое тело настолько плoxo развито, что отграничить их от Hypochnus совершенно невозможно; поэтому здесь нередко наблюдается неустойчивость в названиях. Например, распространенный полупаразит на картофеле и многих других растениях Hypochnus solani называется также Corticium vagum; Hypochnus terrestris называют также Gorticium terrestre.
Рис. 236. Hypochnus subtilis, разрез через сплетение гиф с базидиями.
К Gorticium близки Coniophora, Peniophora и Aleurodiscus, образующиe такие же в общем ресупинатные плодовые тела в виде корочек на мертвых ветвях и на древесине. В частности, Peniophora характеризуется наличием шиповидных цистид, высовывающихся над поверхностью гимения; у Aleurodiscus развиваются между базидиями в гимений сравнительно тонкие парафизы (рис. 237).
Рис. 237. Плодовые тела Aleurodiscus amorphus, справа базидии и парафизы.
По внешности сходны с ним Tulasnella и Vuilleminia. Однако по подробностям строения и развития базидий они нередко выделяются в особую группу. В частности, Tulasnella по своим базидиям, состоящим из гипобазидии и эпибазидии, может сближаться с Tremellales (см. стр. 350—351).
Другой крупный и типичный род Thelephoraceae, Stereum имеет также плоские дорзовентральные плодовые тела, но более массивные, плотные, состоящие из нескольких слоев и нередко по краям приподнимающиеся от субстрата. У некоторых видов плодовые тела приподнимаются и скручиваются в верхней расширенной части в воронку. Они, несут тогда гимений только на ее наружной стороне (рис. 238).
Рис. 238. Плодовое тело Stereum elegans.
При развитии на верхней стороне горизонтального субстрата плодовые тела Stereum ресупинатные с гимением на верхней стороне или отчасти на нижней образуются на приподнимающихся краях; у других видов особенно на вертикальном субстрате образуются плодовые тела, частично отстоящие от субстрата консолевидно. Они прикреплены только одной стороной и разрастаются свободно в горизонтальном положении. Гимений на них развивается только снизу. То же бывает у Hymenochaete, близкой к Slereum (рис. 239).
Рис. 239. a — плодовые тела Hymenochaete tabacina, b — схема расположения гимения (h).
У Thelephora плодовые тела также кожистые, ресупинатные или снабженные ножкой и поднимающиеся более или менее вертикально. В последнем случае они часто рассечены на плоские лопасти или имеют вид воронки с гимением на наружной (морфологически нижней) поверхности. Такое же воронковидное строение имеет Craterellus, плодовые тела которого более или менее мясисты и развиваются прямо на почве. Craterellus весьма приближается к Cantharellus (лисички) также с воронковидными плодовыми телами, но более широко развернутыми в верхней части. Гимений развивается здесь на продольных (радиальных) складках на наружной (нижней) поверхности, весьма сходных с гименофором Agaricaceae (рис. 240).
Рис.240. 1 — Craterellus, 2 — Cantharellus.
Несколько иной путь эволюции намечается у Cyphella и Solenia. Плодовые тела их также вертикальные, воронковидные или трубковидные, но несут гимений на внутренней (морфологически верхней) поверхности. Такое строение намечается у некоторых Aleurodiscus (род близкий к Corticium), где благодаря разрастанию нижних слоев ресупинатное плодовое тело принимает блюдцевидную форму. У Cyphella и Solenia то же развитие пошло еще дальше.
Цитологически, развитие базидий
Изучено у немногих сравнительно представителей Thelephoraceae и оказалось, что здесь встречается и стихобазидиальный, и хиастобазидиальный типы. К первому принадлежат некоторые Craterellus, близкий к нему Cantharellus и, повидимому, Thelephora palmata. У первых двух развитие изучено подробнее: здесь нередко наблюдается и третье деление в базидии, а число спор на ней иногда превышает обычные четыре (рис. 225).
Наоборот, Hypochnus, Corticium, Stereum и др. принадлежат к хиастобазидиальному типу. Таким образом, в семействе Thelephoraceae высота организации, судя по строению плодового тела, с одной стороны, и по развитию базидий, с другой — не совпадает; обстоятельство, которое заставляет некоторых авторов разбивать эту группу. Особо нужно отметить, что Cantharellus по мясистой консистенции плодовых тел и пластинчатому гименофору относится многими систематиками к Agaricaceae, но стихобазидиальный тип базидий, не свойственный Agaricaceae, дает некоторые основания и для отнесения его к Thelephoraceae.
Практическое значение
Craterellus clavatus и Cantharellus cibarius (лисички) — съедобны. Coniophora cerebella — распространенный разрушитель древесных построек (так называемый «погребной гриб»). Stereum hirsutum и S. frustulosum — паразиты на дубе. Последний вызывает очень характерное разрушение его древесины, известное под названием «куропатка» (древесина делается пятнистой, подобно куропатке). Stereum pureum — паразит на плодовых деревьях. Многие другие сапрофитные виды Stereum, Corticium и близких к ним вызывают гниль древесины в лесу и отчасти на складах. Thelephora terrestris (T. laciniata) почвенный гриб, заглушающий иногда плодовыми телами сеянцы хвойных. Hypochnus solani и Н. terrestris — паразиты картофеля и многих других растений (в том числе и сеянцев древесных пород).
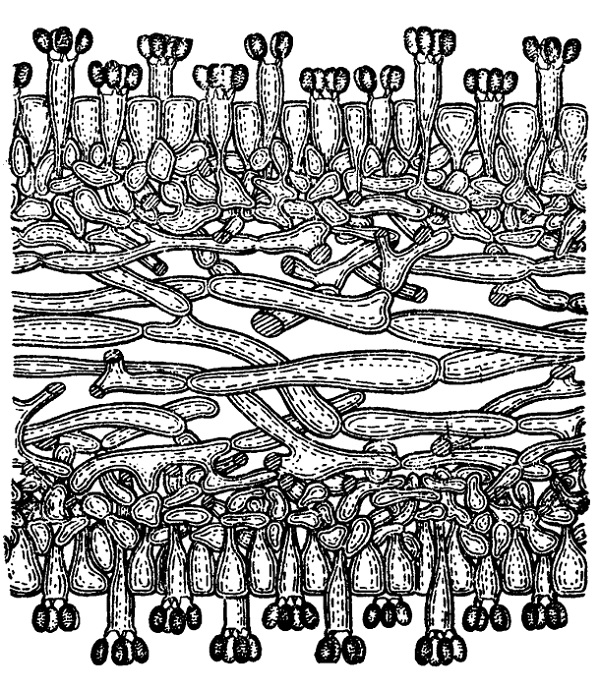
Этот порядок наиболее типичен среди базидиомицетов и вместе с тем наиболее обширен (около 12 тысяч видов). Как показывает название, наиболее характерным признаком считается здесь расположение базидии гимением на поверхности плодового тела. Лишь у немногих простейших представителей гимений зачаточный или совсем не выражен, так же как и плодовые тела. В типичном случае гимений состоит из слоя цилиндрических или слегка расширенных на вершине (булавовидных) базидии и чередующихся с ними парафиз, обыкновенно такой же формы и также в молодости содержащих два ядра, но не сливающихся затем (рис. 235). Реже и преимущественно у простейших представителей парафизы нитевидные, иногда даже ветвящиеся (у видов Aleurodiscus).
Рис. 235. Coprinus comatus: разрез пластинки с гимением; базидии на ней двух сортов: более длинные и более короткие.
У некоторых гименомицетов развиваются еще особые цистиды в виде более крупных, чем парафизы, образований, возвышающихся своими концами, над общим уровнем гимения. Их стенки нередко утолщены или инкрустированы, и они играют главным образом защитную роль, предохраняя молодой гимений от соприкосновения с другими поверхностями, что могло бы повредить развивающиеся базидиоспоры. Что же касается парафиз, то как сочные тургесцентные образования они придают известную упругость всему гимениальному слою и поддерживают с боков базидии, отделяя их вместе с тем друг от друга и предохраняя от слипания сидящие на них базидиоспоры.
Базидии всегда построены по акроспоровому типу, т. е. несут базидиоспоры на своей вершине (почти всегда по 4). Последние по созревании отбрасываются с некоторой силой от поверхности гимения, причем это происходит большей частью не одновременно для всех четырех базидиоспор. После отделения спор базидия отмирает и спадается. Согласно прежним указаниям (Juel, 1898, и др.), там, где как у Cantharellus в базидии в результате трех делений получается 8 ядер и из них 4 переходят в базидиоспоры, а 4 остаются в самой базидии — после отделения первой генерации базидиоспор развивается затем вторая. Однако новейшие исследования (главным образом Buller) не подтверждают этого. Базидия развивает только одну генерацию спор и отмирает вслед за их отделением. У большинства гименомицетов отделение спор с гимения происходит постепенно, так что на одном небольшом участке находятся базидии разного возраста, причем по мере отмирания более старых между ними из более глубоких слоев выдвигаются новые и начинают развивать на своей вершине базидиоспоры. Таким образом, во многих случаях развивается до 4—5 последовательных генераций базидии (например у шампиньонов и многих других Agaricaceae). У некоторых трутовиков гимений функционирует в течение всего вегетационного периода, а у некоторых оказывается даже многолетним (например у Daedalea). Надо думать, что здесь число генераций последовательно образующихся базидии должно быть весьма значительным. Реже встречается другой тип, представленный главным образом у видов Goprinus. Здесь определенный участок гимения функционирует только очень непродолжительное время и после отделения базидиоспор весь отмирает и расплывается. Тем временем тот же процесс переходит на соседний участок и т. д. Однако и у Goprinus с его столь эфемерным гимением удается различить в нем два сорта базидии: более длинные и более короткие. Они развиваются почти одновременно, но все-таки первые раньше сбрасывают свои споры, чем вторые (рис. 235).
Гимений развивается на плодовых телах мясистой, кожистой или деревянистой консистенции. У низших представителей — плодовые тела в виде поверхностного плоского сплетения гиф, неопределенно нарастающего своими краями. Его верхняя свободная поверхность несет гимений, также не ограниченный в своих очертаниях. Такие плодовые тела называются ресупинатными. Они типично представлены, например у Coniophora, Соrticium и некоторых других (из семейства Thelephoraceae). У близкого систематически Stereum такие же плоские плодовые тела обыкновенно имеют приподнимающиеся от субстрата края, и гимений развивается здесь только на их нижней поверхности. Здесь мы видим характерную вообще для гименомицетов ориентировку гимения на нижнюю сторону, что стоит в связи с характером распространения базидиоспор. У относящихся к тому же семейству Thelephora и некоторых других плоские плодовые тела часто стоят почти вертикально, прикрепляясь к субстрату суженным основанием. Гимений здесь — или с обеих сторон или только с одной, обращенной книзу. Наконец, у Crater ellus и некоторых других плодовые тела построены по радиальному типу и имеют вид воронки с узким нижним концом. Гимений находится здесь на наружной, отчасти книзу обращенной поверхности.
Аналогичную эволюцию плодовых тел от ресупинатных плоских до вертикально стоящих и радиальных можно проследить и в других семействах гименомицетов (Hydnaceae, Polyporaceae), причем параллельно с этим изменением формы также наблюдается перенесение гимения на нижнюю сторону.
Другая существенная черта строения плодовых тел гименомицетов заключается в расположении на них гимения. У одних, преимущественно низших форм, несущая гимений поверхность, или гименофор, не выделяется особенно от остальной поверхности плодового тела и оказывается гладкой или слегка бородавчатой (гименофор гладкий). У других гимений располагается на особых выростах плодового тела, имеющих определенную форму (гименофор в виде шипов, трубочек или пластинок). Эти различия кладутся в основу систематического разделения гименомицетов.
Наконец, весьма существенно то, что у одних гимений с самого начала лежит открыто на поверхности плодового тела или его гименофоров, у других же он развивается сначала в замкнутой полости, прикрытый сплетением стерильных гиф, называемых покрывалом (velum), и обнажается лишь при созревании базидий вследствие разрыва или разрушения этого покрывала. Первый тип может быть назван гимнокарпным, а второй гемиангиокарпным. К первому относятся более примитивные группы (Thelephoraceae, Glavariaceae, Hydnaceae и частью Polyporaceae), а второй представлен у высших Polyporaceae (Boletus) и особенно у Agaricaceae. Плодовые тела у этих последних обычно состоят из пенька и шляпки, причем шляпка в молодости прижата свободным краем к пеньку, и здесь между ними существует соединение, образованное покрывалом. Позднее, при развертывании гимения, оно разрывается. У некоторых Agaricaceae, как у Amanita, кроме этого покрывала, называемого частным (velum partiale), имеется еще общее (velum universale), которое в молодости окружает все плодовое тело и разрывается раньше частного покрывала вследствие вытягивания пенька. При этом часть его остается в виде оторочки у основания пенька, а другая часть остается на шляпке в виде лоскутов (такое именно происхождение имеют белые пятна на верхней поверхности шляпки обычного красного мухомора).
Анатомическое строение плодовых тел
При нередко значительных размерах и сложности внешнего расчленения плодовые тела гименомицетов могут достигать также и довольно значительной сложности анатомического строения. Кроющая ткань диференцирована преимущественно на верхней стороне шляпки у Agaricaceae и некоторых Polyporaceae. О строении кроющей ткани см. стр. Кроющая ткань. Нередко она может быть отодрана в виде более темно окрашенной пленки от более светлой и рыхлой внутренней части. Последняя представляет или войлочное сплетение гриф, или в некоторых мясистых плодовых телах носит параплектенхиматический характер. У некоторых Agaricaceae, как у Lactarius, эта внутренняя ткань состоит из гнездами расположенных параплектенхиматических участков и отделяющих их друг от друга гифенных войлочных прослоек. Весьма широко распространены в плодовых телах элементы, проводящие органические вещества (см. рис. 19); у Lactarius и некоторых других они имеют характер млечных трубок, у большинства же просто отличаются более густым содержимым, богатым жиром и другими питательными веществами. Эти проводящие гифы особенно обильно подходят к гимению и иногда высовываются своими концами на поверхность, образуя своеобразные как бы цистиды (глеоцистиды) (рис. 18, 1,2). У трутовиков, как виды Fomes,Trametes и др., плодовые тела отличаются значительной твердостью. Это обусловлено тем, что слагающие их гифы обладают сильно утолщенными прочными стенками, но обыкновенно не замечается ясной группировки этих имеющих механическое значение гиф в определенные механические ткани. Лишь кое-где в пеньках шляпных грибов замечается скопление утолщенных элементов на периферии (кроющая ткань в пеньках большею частью не выражена с достаточной ясностью). Чаще всего упругость и прочность пенька на изгиб обусловлена преимущественно тургором слагающих его гиф и известным напряжением, какое существует между сжатой в продольном направлении внутренней его частью и растянутой наружной. Определенной эмбриональной ткани в плодовых телах гименомицетов не имеется, если не считать за таковую краевой нарастающей зоны у ресупинатных форм. Обычно рост плодового тела обусловлен более или менее равномерным разрастанием всех его частей, хотя иногда отмечается более сильный рост в верхних участках пенька у шляпных грибов (интеркалярная зона роста).
Плодовые тела мясистой консистенции обыкновенно однолетние и часто существуют в развитом состоянии всего несколько дней; что же касается кожистых и особенно деревянистых плодовых тел, то они обычно многолетние. При этом у одних, как Schizophyllum, Daedalea, Trametes и др., все время сохраняется один и тот же гимений, прерывающий только свою жизнедеятельность на время зимы или сухого периода. У других, как представители рода Fomes, гимений функционирует только один сезон, а на следующий год поверх него развивается новый и т. д. Такие плодовые тела, имеющие ясно выраженное зональное нарастание, не только с нижней гимениальной стороны, но и сверху, достигают иногда очень значительных размеров и солидного возраста (до 80 лет).
По образу жизни большинство гименомицетов относится к почвенным макрофитам. Значительное количество их также живет в качестве сапрофитов на древесине или паразитов на стволах деревьев. Общая биология тех и других освещена в 1-й части (см. стр. Древесные грибы, Копрофильные грибы, Почвенные грибы), так же, как и распространение у них спор и соответственно с этим строение и расположение гимения и гименофоров (см. стр. Распространение спор).
Семейства гименомицетов
Порядок Hymenomycetales может быть разделен на следующие семейства:
Гимений расположен непосредственно на поверхности плодового тела (гименофор гладкий). а) Плодовые тела зачаточные, паутинистые или у большинства ясно развитые, кожистой консистенции, большей частью плоские, иногда прямо стоящие, воронковидные. Гимений с одной стороны.
Сем. Thelephoraceae. б) Плодовые тела мясистые, прямо стоящие, булавовидные или ветвящиеся. Гимений большей частью покрывает их со всех сторон.
Сем. Clavariaceae.
Гимений расположен на особых выростах плодового тела (гименофорах). а) Гименофор в виде шипов или зубцов.
Сем. Hydnaceae. б) Гименофор в виде трубок или переплетающихся складок.
Сем. Polyроrасеае. в) Гименофор в виде пластинок, расходящихся радиально от центра (большей частью от пенька).
Сем. Agaricaceae.
Приводимое здесь разделение при всей его внешней простоте и наглядности практически может иногда создать затруднение, так как существуют формы, обнаруживающие промежуточный характер между указанными семействами; поэтому провести резкое разграничение между ними не всегда возможно. С другой стороны, эта система оспаривается некоторыми с точки зрения филогении. Здесь некоторые современные авторы придают преимущественное значение, с одной стороны, строению и развитию базидии (стихо- и хиастобази-диальные), а с другой — гимнокарпности и гемиангиокарпности. Относительно значения первого момента некоторые соображения были высказаны выше (см. стр. Базидиомицеты). Здесь можно только указать, что история развития базидий прослежена у такого сравнительно ничтожного количества форм, что полагать ее в основу систематического разделения всей огромной группы гименомицетов кажется слишком смелым, тем более, что у близких по остальным свойствам видов встречается как тот, так и другой тип развития (например у разных видов Clavaria). Что касается второго, то при всей принципиальной важности его мы также располагаем еще слишком скудными данными относительно истории развития плодовых тел и заложения гимения, чтобы класть этот принцип в основу систематики. Сторонники его, Maire и за ним Гейман, объединяют, например, в свои Agaricales огромное количество видов, которые они считают гемиангиокарпными. При этом сюда относятся не только пластинчатые грибы, но и Boletus. Все остальные считаются гимнокарпными. Однако фактически для огромного большинства тех и других мы совершенно не знаем этой стороны развития. Во всяком случае принятая здесь система имеет преимущество простоты и наглядности, хотя и нельзя отрицать, что, может быть, она не совсем точно иногда отражает филогенетические отношения.
Небольшая группа исключительно паразитов (около 25 видов). Главный род Exobasidium, представителем которого может служить Е. vaccinii, обычный у нас на бруснике и других видах Vaccinium, где вызывает гипертрофии на листьях и молодых стеблях. Гипертрофированная ткань пронизана обильным межклетным мицелием, сложенным из двуядерных клеток. Образующиеся на нем базидии высовываются между клетками эпидермиса, образуя на пораженном органе беловатый налет, но не слагаются в сомкнутый слой (гимений) (рис. 234).
Рис. 234. Exobasidium vaccinii: a — внешний вид пораженной ветви брусники; b — разрез через лист, базидии просовываются между клетками эпидермиса.
При развитии базидии два первоначальных ядра ее сливаются, и копуляционное ядро делится, причем ось деления располагается вдоль базидии (стихобазидиальный тип). Сколько происходит здесь всего делений, два или три, с точностью не установлено, однако, судя по тому, что зрелая базидия несет от двух до шести спор (всего чаще 4), более вероятным кажется последнее. При прорастании базидиоспоры делятся на 2 — 4 клетки, каждая из которых развивает короткую нить и на ней конидии. Последние могут размножаться почкованием, и затем вырастают в мицелий. Exobasidium rhododendri вызывает на листьях Rhododendron образование крупных мясистых разрастаний (галлов) красноватого цвета.
У тропического Kordyana базидии, залагаясь в дыхательной полости пораженного листа, высовываются небольшими пучками через устьица вместе с перепутанными стерильными гифами. Число базидиоспор здесь также не строго фиксировано, но в большинстве случаев их две.
Небольшая группа (около 30 видов) сапрофитных грибов с студенистыми, чаще всего ярко окрашенными плодовыми телами различных внешних очертаний: в виде корочек (Сегасеа), полушаровидных морщинистых подушечек (Dacryomyces), цилиндрических, коралловидных разветвленных тел (Calocera) и т. п. У Dacryomitra, плодовые тела расчленены на пенек и морщинистую шапочку и весьма напоминают миниатюрные сморчки (рис. 232).
Со стороны истории развития изучен Dacryomyces deliquescens, нередка встречающийся на гнилой древесине.
Базидиоспоры здесь, как это типично для всей группы, перед прорастанием разделяются на несколько клеток. На них образуются многочисленные мелкие конидии, позднее вырастающие в мицелии. Плодовые тела в виде полушаровидных подушечек сначала отчленяют многочисленные оидии, а затем развивают базидии. Последние имеют цилиндрическую форму и расположены палисадным слоем по поверхности плодового тела под общим слоем одевающей его слизи (рис. 233).
Рис. 233. Dacryomyces deliquescens, разрез плодового тела с базидиями.
Как и все клетки плодового тела, базидии первоначально двуядерны, затем в них происходит кариогамия, и копуляционное ядро дважды делится, причем оси делений располагаются вдоль базидии (стихобазидиальный тип). Тем временем на верхнем конце ее образуется два толстых выроста, которые, Достигнув поверхности общей слизи, развивают на концах по одной тонкой стеригме и на них по одной базидиоспоре. В базидиоспоры переходит по одному ядру, а два другие остаются в базидии и там дегенерируют.
Представители Dacryomycetales встречаются преимущественно на гнилой древесине. Практического значения они не имеют. О филогенезе Dacryomycetales см. ниже на стр. 350.
К базидиомицетам относится более 20 тысяч видов. Подобно тому как другая большая, группа высших грибов, аскомицеты, характеризуется наличием сумки как основного спороношения, так базидиомицеты характеризуются базидией. Последняя развивается на диплоидном мицелии и в молодости содержит, как и сумка, два ядра (дикарион), которые затем сливаются. Выше уже было указано на гомологию сумки и базидии (стр. Введение в курс проф. Л. И. Курсанова «Микология»). В соответствии с этим и эволюция обеих указанных групп, происходящая на основе гомологичных спороношений, показывает значительные черты параллелизма. Основные различия между аскомицетами и базидиомицетами могут быть вкратце сформулированы следующим образом:
У аскомицетов морфологически основное спороношение (сумка) образует эндогенные споры (аскоспоры), а у базидиомицетов морфологически основное спороношение (базидия) образует экзогенные споры (базидиоспоры).
У аскомицетон сумки развиваются в типе в результате оплодотворения в виде слияния особо диференцированных половых органов, хотя у многих настоящее оплодотворение утрачивается и заменяется различными формами апогамии. У базидиомицетов первоначальное оплодотворение и половые органы полностью утрачены, и развитие идет исключительно апогамно.
У аскомицетов, как правило, в цикле развития преобладает гаплоидная фаза, представленная вегетативным мицелием, различными конидиальными спороношениями на нем и главной массой сплетения гиф, образующих аскокарп. Диплоидная фаза у них представлена только аскогенными гифами, заключенными в ткани плодового тела и питающимися на счет гаплоидной ткани последнего. У базидиомицетов гаплоидная и диплоидная фазы представлены в виде двух самостоятельных мицелиев. При этом более развита диплоидная фаза, представленная мощно развитым вегетативным мицелием, иногда конидиальными спороношениями на нем и всем базидиальным плодовым телом (отличие от аскокарпа сумчатых грибов, где плодовое тело в большей своей части гаплоидно). Что касается гаплоидной фазы базидиомицетов, то она в типе представлена в виде особого мицелия, но развита значительно слабее, а во многих случаях сильно сокращена и даже полностью выпадает (гаплоидными остаются только молодые базидиоспоры).
Развитие базидии происходит следующим образом. Она берет начало из клетки с двумя ядрами (дикарион), занимающей вершину такой же дикариофитной (с двуядерными клетками) гифы. Нужно сказать, что у большинства базидиомицетов такие диплоидные дикариофитные гифы, кроме особенности своего ядерного аппарата, отличаются еще наличием пряжек — особых маленьких клеток, лежащих сбоку против поперечных перегородок (рис. 221).
Рис. 221. Развитие базидии гименомицета: 1 и 2 — материнская клетка базидии; 3 — одновременное деление ее двух ядер, одно из дочерних попадает в зачаток пряжки; 4 — верхняя клетка — молодая базидия — содержит два ядра, клетки пряжки и ножка — по одному ядру; 5 — переход ядра из пряжки в клетку ножки; 6 — два ядра базидии слились в одно; 7 — копуляционное ядро дважды разделилось, на вершине базидии — зачатки стеригм.
На рис. 221, 1 как раз изображено начало такой пряжки в виде небольшого бокового выроста. Вскоре после его образования оба ядра дикариона одновременно делятся, причем оси делений лежат на одном уровне и вдоль клетки (рис. 221, 3). Из получившихся четырех ядер два отходят кверху, одно попадает в вырост пряжки и одно отходит в нижнюю часть клетки. Затем делится и самая клетка поперечной перегородкой, расположенной немного ниже отхождения зачатка упомянутой пряжки. В результате этого верхняя клетка, представляющая материнскую клетку базидии, получает два несестринских ядра, или дикарион, нижняя же клетка, или клетка-ножка, получает только одно ядро, так как второе попадает в вырост пряжки (рис. 221, 4). В дальнейшем этот вырост отделяется перегородкой от основания материнской клетки базидии; вершина его, загибаясь книзу, прирастает к клетке-ножке и через образующееся отверстие отдает ей свое ядро (рис. 221, 5). Благодаря этому клетка-ножка восстанавливает двуядерность, свойственную диплоидной фазе, и делается способной к дальнейшему развитию, прорастая в новую ветвь (рис. 221, 5) с таким же развитием базидии на ней. Что касается материнской клетки базидии, то ее дикарпон сливается в одно диплоидное ядро (рис. 221, 6), и последнее делится обыкновенно дважды, причем происходит редукция числа хромозом, так что в молодой базидии получаются четыре гаплоидных ядра (рис. 221 ,7). В это время на вершине базидии появляются четыре выроста, которые, вздуваясь на конце, развивают каждый по базидиоспоре. В каждую из них через узкий канал ножки или стеригмы протискивается по одному гаплоидному ядру. По созревании базидио-споры с известной силой отбрасываются от своих стеригм (см. стр. Распространение спор).
Если сравнить эту историю развития базидии с тем, что было указано относительно сумки, то получится значительное сходство. Кончик крючка аскогенной гифы соответствует пряжке. Они сходны не только по положению, но и по тому, что получают тот и другая по одному ядру и отдают его клетке-ножке. Также сходны материнские клетки сумки и базидии. Они получают по два ядра (дикариона), которые сливаются в одно диплоидное, и последнее делится затем редукционным путем (рис. 222).
Рис. 222. Параллелизм крючков на аскогенных гифах с развивающимися на них сумками (1— 6) и пряжек диплоидного мицелия с базидиями (7—12); схема.
Правда, в сумке большей частью происходит три деления и получается восемь гаплоидных ядер, а в базидии только четыре, но это различие не строго постоянно, так как известны сумки, в которых имеется только два деления ядра, и базидии, в которых происходит три деления. Единственное постоянное отличие состоит в том, что аскоспоры образуются эндогенно, а базидиоспоры экзогенно, как это видно из схемы на рис. 223.
Рис. 223. Схема развития сумки (верхний ряд) и базидии (нижний ряд).
В подробностях строение и развитие базидии может быть различным.
Прежде всего различают холобазидии и фрагмобазидии. В первом случае базидия остается неразделенной одноклеточной, а во втором вслед за делением копуляционного ядра и сама она разделяется или поперечными перегородками на четыре друг за другом расположенные одноядерные клетки, или продольными перегородками на четыре клетки, расположенные на одном уровне. Каждая из этих четырех клеток развивает уже только по одной базидиоспоре (рис. 224).
Рис. 224. Базйдии: 1 — холобазидии: а — Coprinus (акроспоровая); 2 — Tulostoma (плевроспоровая); с — Dacryomyces (двуспоровая); b — фрагмобазидии: а — Tremella (вдоль разделенная); b — Endophyllum (поперек разделенная склеробазидия).
Кроме указанного различия, холобазидии и фрагмобазидии отличаются еще и ходом своего развития. Первые формируются полностью из производящей их материнской клетки. В ней происходит кариогамия, деление копуляционного ядра и, наконец, на вершине образуются только тонкие стеригмы, несущие на концах базидиоспоры. Фрагмобазидии имеют более сложное развитие. В них можно различить две части: гипобазидию и эпибазидию. Первая, как, и холобазидия, формируется прямо из материнской клетки. В ней также происходит кариогамия, а часто также и деление копуляционного ядра. Что же касается эпибазидии, то она представляет вторичный вырост на гипобазидии, отсутствующий в типичных холобазидиях. Лишь на концах этих выростов образуются тонкие стеригмы с базидиоспорами. Параллельно с этим в большинстве случаев происходит деление всей базидии, т. е. она делается фрагмобазидией. Однако последнее происходит не всегда. Например, у Dacryomycetales две толстые ветви на вершине цилиндрической базидии представляют по развитию эпибазидию, однако, деления на клетки здесь не происходит, так что формально все образование может называться холобазидией, хотя и отличается от нее по своему развитию (рис. 224, с). Ввиду этого предлагается иногда вместо термина фрагмобазидии другой — гетеробазидия, охватывающий все случаи с указанным более сложным развитием, независимо от того, останется ли в конце концов все образование одноклеточным (как у Dacryomycetales) или делается многоклеточным (как у Tremellales и др.).
Указанные различия в строении и развитии базидии имеют наибольшее значение и кладутся в основу для разделения всего класса грибов на два подкласса. В свою очередь они могут формулироваться несколько различно: Нolobasidiomycetes и Phragmobasidiomycetes, если в основу полагается формальный признак одноклетности или многодетности базидии. Если же за основу взять историю развития базидии и отсутствие или наличие эпибазидии, то название второго подкласса может быть заменено на другое: Heterobasidiomycetes. При этом объем его оказывается несколько шире, чем Phragmobasidiomycetes (в него входят и формы с неразделенной базидией, как у Dacryomycetales). Хотя второй принцип, повидимому, более отражает филогенетические отношения, но практически чаще применяется первый, т. е. производят деление базидиомицетов на Holobasidiomycetes и Phragmobasidiomycetes, как более наглядное.
Менее существенно расположение базидиоспор. У большинства представителей они располагаются на вершине базидии на одном уровне. Такие базидии называются акроспоровыми. Они типичны для большинства холобазидии, особенно у гименомицетов, где базигоспоры по созревании активно отбрасываются, а также для некоторых фрагмобазидий, разделенных продольными перегородками (у Tremellales, рис. 224, 2а). В других случаях базидиоспоры располагаются по бокам базидии на разных уровнях. Такие базидии называются плевроспоровыми. Они встречаются у немногих холобазидиомицетов, с замкнутыми плодовыми телами, где активного отбрасывания базидиоспор не имеется (рис. 224, 1 b), а также у тех фрагмобазидиомицетов, базидия которых разделена на клетки, расположенные друг над другом (рис. 224, 2 b).
Число базидиоспор — четыре — также типично для базидии, как восемь аскоспор для сумок. Однако из этого правила известно немало исключений. Известны односпоровые базидии (у Pistillaria maculicola); значительно чаще встречаются двуспоровые (например у культурного шампиньона или у Dacryomycetales (рис. 224, 1 с). Наконец, известны базидии с числом спор больше чем четыре. Например, у Cantharellus cibarius обыкновенно шесть спор на базидии, у Corticium coronatum, Tilletia tritici — восемь, а у близкого к Tilletia другого представителя головневых, Neovossia, — их несколько десятков (см. рис. 277, 5).
В случаях двуспоровых базидий установлено, что при их развитии копуляционное ядро большей частью делится, как обычно, два раза и даже иногда три раза (например у Clavaria cristata и Cl. rugosa). Дальнейшая судьба получившихся четырех или даже восьми гаплоидных ядер в базидии различна в разных случаях. У двуспоровых видов Glavaria, Stilbumcarneum и Dacryomycetales два ядра переходят по одному в каждую спору, а остальные дегенерируют в базидии. У Hydnangium carrieum и культурного двуспорового шампиньона каждая базидиоспора получает по два ядра. Такие первично двуядерные споры в некоторых случаях оказываются гомоталличными в отличие от гетероталличных четырехспоровых рас, где каждая спора получает по одному ядру. Например, это установлено у Naucoria semiglobata и Galera tenera (Sass, 1929). Аналогичное явление наблюдается и у сумчатых грибов, например, у четырехспоровых и восьмиспоровых Neurospora (см. стр. Гетероталлизм).
В случаях многоспоровых базидий, конечно, должно быть больше двух делений копуляционного ядра, что фактически установлено, например, у Tilletia, где копуляционное ядро делится три и даже четыре раза, образуя до 16 ядер в базидии. Следует отметить, что у некоторых базидиомицетов, имеющих обычно четырехспоровые базидии, тем не менее в них образуется восемь ядер, из которых далее функционируют только четыре (Gantharellus, некоторые Clavaria и др.).
Далее, имеются различия в самом расположении ядерных делений в развивающейся базидии. У большинства оси первого и большей части второго деления расположены поперек, а у некоторых вдоль базидии. Первые называются хиастобазидиальными, а вторые — стихобазидиальными (рис. 225).
Многие стихобазидиальные базидиомицеты имеют вместе с тем третье деление ядер в базидии, что напоминает нормальное развитие сумки. Поэтому некоторые авторы (Juel, Maire, Gaumann) рассматривают этот тип как примитивный, объединяя имеющие его виды (различного внешнего строения) в группу Cantharellales, из которой уже выводятся более высоко диференцированные группы.
Наконец, последней особенностью, которую здесь надлежит отметить, является то, что в некоторых случаях базидия развивается из толстостенной клетки, имеющей большей частью значение покоящейся споры. Базидии этого последнего типа называются склеробaзидиями. Они типичны для некоторых групп фрагмобазидиомицетов (Ustilaginales, Uredinales).
Базидиоспоры всегда одноклетны и большей частью одноядерны. Лишь у немногих представителей они оказываются первично двуядерными, как это было указано выше. Более часто встречается вторичная двуядерность базидиоспор, когда последние, будучи сначала одноядерными, делаются затем, еще сидя на базидии, двуядерными вследствие деления первоначального ядра. Это наблюдается, например, у многих гастеромицетов или у Hypochnus terrestris.
По форме базидиоспоры чаще всего округлые или овальные, редко нитевидно вытянутые, как у Tilletia и некоторых других. Оболочка их в одних случаях бывает тонкой и бесцветной, в других — окрашенной, более толстой и с различной «скульптурой» на поверхности в виде шипиков и т. п. Окрашенная оболочка часто состоит из двух слоев: наружного (экзоспорий), содержащего пигмент, и внутреннего (эпдоспорий) бесцветного. У некоторых представителей с двойной споровой оболочкой различается особая ростковая пора. Она расположена на дистальном конце споры и образуется вследствие недоразвития в этом месте экзоспория (например у шампиньона).
Прорастают базидиоспоры большей частью прямо мицелием, реже почкованием или образованием вторичных конидий (рис. 272, 2), которые лишь позднее развивают нитчатый мицелий.
Рис. 272. Auricularia auricula judae: 1 — разрез поверхностного слоя плодового тела; 2 — базидиоспоры и их прорастание.
У некоторых простейших базидиомицетов базидиоспоры при прорастании получают перегородки, делящие их на несколько клеток. Из них каждая прорастает, развивая обыкновенно вторичные почкующиеся конидии. Такой способ прорастания базидиоспор характерен для Dacryomycetales, Tremellales и Auriculariales, которые даже объединяются иногда по этому признаку в особую группу Phragmosporeae.
Так как базидиоспора содержит гаплоидное ядро, получившееся в результате редукционного деления в базидии, то она начинает гаплоидную генерацию, в виде вырастающего из нее гаплоидного или, как здесь нередко говорят, первичного мицелия. Цитологически он характеризуется в некоторых случаях тем, что слагающие его клетки одноядерны (например у ржавчинных грибов). В других случаях, например у многих гименомицетов, развитие гаплоидного мицелия происходит несколько иначе. Базидиоспора при прорастании делается многоядерной, и из нее вырастает многоядерная ветвящаяся гифа, которая растет дальше, не получая некоторое время поперечных перегородок (рис. 226).
Рис. 226. Прорастание базидиоспоры Coprinus с образованием многоядерных гиф, не разделенных на клетки.
Затем такие перегородки появляются, разделяя мицелий на многоядерные клетки. У многих гименомицетов в таком многоядерном состоянии гаплоидный мицелий продолжает развиваться и дальше. Он может размножаться бесполым путем при посредстве оидий или мелких приподнимающихся на коротких ветвях конидий (рис. 227). Как те, так и другие содержат обыкновенно только по одному ядру, обнаруживая этим свою гаплоидную природу.
Рис. 227. 1 — Coprinus fimetarius, часть гаплоидной гифы с оидиями; 2 — а, b, с — Corticium, диплоидные гифы с пряжками; 3 — Corticium serum, возникновение диплоидной клетки (с зачатками пряжки) из места анастомоза двух молодых гаплоидных гиф.
Гаплоидный мицелий базидиомицетов имеет обыкновенно непродолжительное существование, и на нем развивается затем диплоидный, или вторичный, мицелий. Последний цитологически характеризуется во многих случаях двуядерностью слагающих его клеток (например у ржавчинных грибов и некоторых гименомицетов), в других же случаях клетки его многоядерны, но ядра их ассоциированы в дикарионы, что обнаруживается в момент их деления, когда ядра сближаются попарно и делятся одновременно и параллельно друг другу.
У гомоталличных базидиомицетов переход от гаплоидной к диплоидной фазе осуществляется в пределах одного мицелия, а у гетероталличных — при соединении двух разнородных в половом отношении гаплоидных мицелиев. Внутренние процессы, происходящие при этом, выяснены далёко не достаточно. Относительно гомоталличных форм можно только предполагать, что здесь, вероятно, происходит переход ядра из одной клетки в другую на месте очень обычных для такого первичного мицелия анастомозов между ветвями его гиф. Возможно также, что в некоторых случаях первые дикарионы образуются в результате перехода ядра из одной клетки гифы в рядом расположенную соседнюю или даже просто в результате деления одного ядра клетки без последующего появления между ними перегородки. Относительно гетероталличных форм имеется больше непосредственных наблюдений в этом отношении. В основном здесь имеют значение анастомозы между ветвями двух различных мицелиев, через которые и происходит переход ядра из одной клетки в другую. У некоторых видов диплоидный мицелий начинается прямо от места анастомоза. Получившаяся в этом месте первая двуядерная клетка разрастается дальше, причем оба ее ядра ведут себя как дикарион, делясь одновременно и параллельно друг другу. В результате этого из первой двуядерной клетки развивается диплоидный мицелий с дикарионами (рис. 228).
Рис 228. Схема происхождения диплоидного мицелия гименомицета из анастомоза двух гаплоидных.
В других случаях диплоидный мицелий возникает на известном, иногда значительном расстоянии от места анастомоза. В опытах Беллера появление диплоидных гиф наблюдалось на расстоянии нескольких сантиметров от того места, где соединились два мицелия (см. стр. Диплоидизация мицелия). Надо думать, что в этих случаях ядро одного пола, перешедшее в клетку другого пола, не ассоциируется тут же с ее ядром в первый дикарион, а переходит через отверстия, образующиеся в поперечных перегородках гифы, в соседние клетки и, перемещаясь таким образом все дальше и дальше, где-нибудь в отдаленной клетке ассоциируется в дикарион с ядром другого пола. Весьма вероятно, что такое диплоидизирующее ядро во время указанного пути неоднократно делится, так что в конце концов может получиться несколько точек диплоидизации. Беллер смог даже определить скорость перемещения диплоидизирующего ядра вдоль гиф. В его опытах она оказалась около 4,5 мм в 1 час (см. стр. Диплоидизация мицелия).
Кроме непосредственного срастания двух разнородных в половом отношении гаплоидных мицелиев, имеется и другой путь диплоидизации у гетероталличных— базидиомицетов: перенесение гаплоидных оидий или конидий с мицелия одного пола на мицелий другого пола. Этот путь имеет основное значение у ржавчинных грибов, а также играет известную роль и у некоторых гименомицетов (см. стр. Диплоидизация мицелия). На схемах, изображенных на рис. 229 и 230, еще раз иллюстрируется переход от гаплоидной к диплоидной фазе у гетероталличных базидиальных грибов. На рис. 229, 1 изображены две базидиоспоры с черным и светлым ядром (+ и — ). Из каждой развивается короткий мицелий, и между ними получается анастомоз с образованием первого дикариона (рис. 229, 3). Далее представлено развитие из этой первой двуядерной клетки многоклетной диплоидной гифы с пряжками (рис. 229, 4—7). Наконец, на конечных разветвлениях ее изображено развитие базидий (рис. 229, 8).
Рис. 229. Схема развития гетероталличного базидиомицета: 1 и 2 — базидиоспоры разных полов (черное и светлое ядро) и их прорастание; 3 — анастомоз; 4, 5, 6, 7 — развитие из анастомоза диплоидной дикариофитной гифы; 8 — развитие базидий на концах диплоидной гифы.
Схема рисунка 230 поясняет аналогичный процесс у гименомицета с шляпным плодовым телом. На его базидиях образуются базидиоспоры (+ и —), прорастающие каждая в гаплоидный мицелий (светлый). Из анастомоза их развивается диплоидный мицелий (темный) и на нем также полностью диплоидное плодовое тело (темное).
Рис. 230. Схема цикла развития шляпного гриба: гаплоидная фаза светлая, диплоидная — покрыта точками.
В отличие от аскомицетов, где диплоидная фаза представлена аскогенными гифами, не имеющими самостоятельного существования, у базидиомицетов диплоидный мицелий существует самостоятельно и преобладает в цикле развития, являясь у многих форм даже многолетним (например у многих трутовиков или почвенных гименомицетов). Далее, диплоидный мицелий базидиомицетов нередко получает значительную морфологическую диференцировку в виде образования ризоморф и особенно шнуров и тяжей, столь распространенных вблизи плодовых тел многих почвенных гименомицетов и гастеромицетов. Наоборот, гаплоидный, или первичный, мицелий развит в большинстве случаев слабо. Обычно, наблюдая в естественной обстановке мицелий базидиального гриба, мы имеем дело с диплоидным его состоянием.
Иногда, как у Hypochnus terrestris, последний совсем отсутствует, так как там двуядерная базидиоспора прямо прорастает в двуядерный же диплоидный мицелий. У Ustilaginales базидиоспоры одноядерны, но при прорастании они часто сразу копулируют друг с другом, и из них прямо вырастает диплоидный мицелий. Таким образом, и здесь гаплоидный мицелий фактически отсутствует. У многих видов гименомицетов могут анастомозировать и давать диплоидный мицелий очень молодые проростки (гаплоидные) базидиоспор. С другой стороны, у некоторых эта способность получается только после некоторого периода роста. Часто момент наступления диплоидности определяется внешними условиями: не только встречей двух подходящих мицелиев ( + и —) у гетероталличных форм, но и условиями культуры: например, погружение мицелия в жидкость задерживает появление диплоидности (Kniep, 1919).
Характерную морфологическую особенность диплоидного мицелия многих базидиомицетов представляют пряжки — маленькие полукруглые клетки, лежащие сбоку на гифе против поперечной перегородки в ней (рис. 227, 2). Развитие их происходит сходно с тем, что было указано выше, при описании формирования базидии (см. выше на этой странице). При делении ядра дикарионы располагаются рядом посредине клетки, и против них образуется короткий боковой вырост, загибающийся вниз по направлению к основанию гифы. Когда ядра дикариона разделяются, то из четырех вновь образовавшихся ядер два отходят в верхний (дистальный) конец клетки, одно в нижний ее конец (проксимальный) и одно в упомянутый вырост, являющийся зачатком пряжки. Затем появляется поперечная перегородка как раз под основанием выроста. Ею отделяется верхняя двуядерная клетка гифы от нижней одноядерной. Вскоре появляется перегородка в основании зачатка пряжки, а затем появляется отверстие между дистальной частью нижней клетки гифы и прилегающим к ней концом пряжки. Через это отверстие ядро последней переходит в нижнюю клетку гифы и восстанавливает таким образом ее двуядерность. У некоторых видов, например у Merulius lacrymans, пряжки часто прорастают в боковую ветвь. Как обстоит дело с ядрами в таких случаях, не выяснено. Какое значение имеет такое сложное перемещение ядра — не известно, морфологически же пряжки соответствуют крючкам, образуемым на аскогенных гифах аскомицетов; они также обычно соединяются с одноядерной первоначальной клеткой, лежащей ниже, и навливают ее двуядерность. О соответствии крючков и пряжек дает ясное представление рисунок 222, изображающий параллельно развитие аска и базидии. Наблюдаемый здесь полный параллелизм вместе с тем особенно резко подчеркивает гомологию того и другого органа.
Пряжки встречаются почти во всех группах базидиомицетов, но особенно распространены среди гименомицетов. Однако и здесь известно некоторое количество форм, которые не имеют пряжек на диплоидном мицелии (например опенок или шампиньон и др.). Таким образом, если присутствие пряжек может служить достаточным указанием на диплоидность мицелия, то отсутствие их не может иметь абсолютно противоположного значения, т. е. указания на гаплоидность его. Это относится даже к гименомицетам, не говоря уже о других группах базидиальных грибов, где пряжки вообще встречаются изолированно только у некоторых видов.
Диплоидный мицелий, как и гаплоидный, может размножаться бесполым путем: оидиями или конидиями, причем они содержат здесь (на диплоидном мицелии) обыкновенно по два ядра и вырастают снова в диплоидный мицелий. Однако в некоторых случаях наблюдалось образование более мелких одноядерных конидий или оидий или разделение первоначальной двуядерной конидии на две одноядерные клетки (например Pholiota aurivella, Martens, 1932). В этом случае такие одноядерные зачатки прорастают в гаплоидный мицелий, т. е. происходит обратный переход от диплоидной к гаплоидной фазе, притом без редукционного деления, вегетативным путем.
Бесполое размножение посредством конидий сравнительно мало распространено как в недолговечной обыкновенно гаплоидной фазе, так и в длительной диплоидной. Исключением являются здесь ржавчинные грибы с обильным развитием конидий и в той и другой фазе, или Fomes annosus, развивающий на диплоидном мицелии обильные конидии на конидиеносцах типа Oedocephalum (рис. 231).
Иное дело оидии или приближающиеся к ним конидии, образующиеся цепочками на мало диференцированных веточках мицелия (рис. 227, 1). Они очень распространены у базидиальных грибов как на гаплоидном, так и на диплоидном мицелии и нередко играют большую роль в их размножении, особенно ввиду того, что и сами могут неограниченно размножаться в питательной среде посредством почкования (например у головневых грибов).
На диплоидном мицелии образуются базидии. У некоторых представителей они развиваются непосредственно на мицелии или берут начало из спор, в свою очередь развившихся прямо на мицелии (склеробазидии головневых и ржавчинных). У большинства базидии образуются на особых плодовых телах, аналогичных плодовым телам аскомицетов, но не гомологичных им: они здесь нацело образованы диплоидными гифами, а не состоят, как у аскомицетов, в главной массе из гаплоидного сплетения, содержащего только незначительные по объему диплоидные части в виде аскогенных гиф. В некоторых случаях отмечается развитие у базидиомицетов гаплоидных плодовых тел, возникающих на односпоровом мицелии у гетероталличных видов. Они сложены из одноядерных клеток, и развивающиеся на них базидии содержат с самого начала одно ядро, делящееся затем два раза (конечно, без редукции хромозом).
В простейшем случае плодовые тела у базидиомицетов представляют плоское гифенное сплетение на субстрате, нарастающее своими краями. Такие плодовые тела неопределенных размеров и очертаний, несущие базидии на своей верхней свободной поверхности, носят название ресупинатных (например у Gorticium, Merulius).
У более высоко организованных представителей плодовые тела, сохраняя плоскую форму, отчасти приподнимаются от субстрата и переносят образование базидий на нижнюю сторону. Дальнейший шаг эволюции представляют трутовики с их копытообразными плодовыми телами, боком приросшими к субстрату и несущими базидии на нижней стороне. Наивысший тип развития в этом направлении представляют прямо стоящие плодовые тела шляпных грибов, у которых базидии образуются также только с нижней стороны шляпки. Перенесение базидий на нижнюю сторону объясняется характером рассеивания базидиоспор (см. стр. Распространение спор) и отчасти, может быть, тем, что в этом положении они оказываются более защищенными от внешних влияний (дождь, высыхание и т. п.). У высших представителей гименомицетов (из сем. Agaricaceae) базидии залагаются не свободно на поверхности, а прикрытые наружным сплетением гиф; лишь при созревании они освобождаются вследствие разрушения или разрыва прикрывающего слоя. Такие плодовые тела называются гемиангиокарпными в противоположность гимнокарпным, где базидии развиваются открыто на поверхности их. От гемиангиокарпных плодовых тел один шаг к ангиокарпным, т. е. вполне замкнутым, характерным для гастеромицетов. У них базидии образуются внутри плодового тела и совсем не обнажаются. Лишь после отделения ими базидиоспор последние освобождаются благодаря разрыву или разрушению общей плодовой оболочки.
Систематическое разделение базидиомицетов
Основывается, во-первых, на строении базидии, ее одноклетности или многоклетности и затем на строении плодового тела. Таким образом, устанавливаются прежде всего два подкласса: Holobasidiomycetes с одноклетными базидиями и Phragmobasidiomycetes с многоклетными базидиями. Первые часто называются также Autobasidiomycetes, а вторые— Protobasidiomycetes. Эта вторая терминология, берущая начало от Брефельда, выражает вместе с тем его представление об эволюции базидиальных грибов: Protobasidiomycetes рассматривается как примитивная группа, от которой берет начало высшая — Autobasidiomycetes. Действительно, представители первой группы никогда не достигают той высоты организации, какая замечается у второй, поэтому представление о Protobasidiomycetes как о примитивной группе сохранилось до настоящего времени. Однако, признавая гомологию базидии с сумкой, нужно принять, что неразделенная базидия (холобазидия или автобазидия) гораздо ближе стоит к сумке, чем разделенная (фрагмобазидия или протобазидия). Что же касается кажущейся большей простоты фрагмобазидиомицетов, то необходимо принять во внимание, что большинство их, притом наиболее типичных (Ustilaginales и Uredinales) живет в условиях паразитизма, не способствующих выработке сложных плодовых тел. Поэтому название Protobasidiomycetes должно быть оставлено и заменено нейтральным в смысле филогенетической значимости названием Phragmobasidiomycetes. Их нужно, вероятно, рассматривать как боковую ветвь от основного ствола Holobasidiomycetes, берущую свое начало от низших его представителей и эволюционировавшую в сторону приспособления к паразитизму и утраты плодового тела.
Систематическое деление базидиомицетов
Базидиомицеты могут быть разделены на следующие основные группы. А. Базидии одноклетные (холобазидии) — подкласс Holobasidiomycetes. I. Базидии вильчато-разветвленные. Базидиоспоры при прорастании делятся на несколько клеток. Сапрофиты, с плодовыми телами студенистой консистенции. 1. Порядок Dacryomycetales. II. Базидии не разветвленные. 1. Плодовых тел не образуется. Базидии развиваются прямо на мицелии одиночно или рыхлым слоем. Базидиоспоры при прорастании делятся на несколько клеток. 2. Порядок Exobasidiales. 2. Базидии развиваются на плодовых телах. а) Базидии располагаются тесным слоем (гимением) открыто на поверхности плодового тела (гимнокарпные) или полузакрыто (гемиангиокарпные). 3. Порядок Hymenomycetales. б) Базидии расположены внутри плодового тела (ангиокарпные). 4. Порядок Gasteromycetales.
Б. Базидии многоклетные (фрагмобазидии)— подкласс Phragmobasidiomycetes. I. Сапрофиты или полупаразиты, образующие плодовые тела студенистой консистенции. 1. Базидии разделены поперечными перегородками на 4 друг над другом лежащие клетки. 5. Порядок Auriculariales. 2. Базидии разделены продольными перегородками на 4 рядом лежащие клетки. 6. Порядок Tremellales. II. Паразиты. Плодовых тел нет. Базидии развиваются из покоящихся спор (склеробазидии). 1. Базидии по строению и по числу клеток не вполне определенные. Они развиваются из хламидоспор, образующихся на мицелии интеркалярно в глубине ткани пораженного растения. 7. Порядок Ustilaginales. 2. Базидии из четырех клеток, расположенных друг над другом. Они развиваются из телейтоспор, образующихся из конечных разветвлений мицелия. Кроме телейтоспор имеются обильные конидиальные спороношения как на Гаплоидном, так и на диплоидном мицелии. 8. Порядок Uredinales.
Дискомицеты, морфологически характеризуемые строением плодовых тел, развитых в виде апотеция, вместе с тем филогенетически не представляют, повидимому, вполне однородной группы. В этом отношении они сходны с пиреномицетами, у которых мы также видим довольно четкую морфологическую характеристику при отсутствии полного филогенетического единства (см. стр. Pyrenomycetes (Пиреномицеты)).
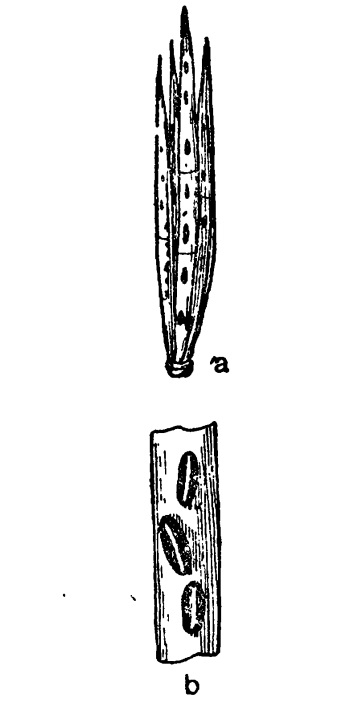
В общем дискомицеты можно рассматривать как дальнейший и наивысший этап эволюции сумчатых грибов, выражающийся наиболее совершенным приспособлением к активному рассеянию аскоспор по мере их созревания. В связи с этим у них выработан гимений, расположенный широким слоем открыто на поверхности плодового тела и ориентированный вверх. Его плотное сложение из узких цилиндрических сумок и нитевидных парафиз между ними обеспечивает, с одной стороны, максимальную продукцию спор на данной поверхности, а с другой стороны, — их активное выбрасывание по мере созревания, так как парафизы поддерживают с боков оболочку сумки, не давая ей раздуваться от возрастающего тургорного давления. Благодаря этому вся сила последнего направлена на верхний свободный конец сумки, где помещаются аскоспоры, всплывающие как более легкие тела (содержание в них масла) в жидком содержимом сумки. При наступающем, наконец, разрыве сумковой оболочки на вершине содержимое сумки с силой выпрыскивается, и аскоспоры подбрасываются кверху на 10 см и даже более. После этого пустая сумковая оболочка спадается, и на ее место начинают врастать снизу новые сумки. Благодаря такой повторности их образования, конечно, значительно увеличивается продукция аскоспор на данной поверхности. С этой стороны дискомицеты, часто отличающиеся также и более крупными размерами плодовых тел, далеко превосходят других аскомицетов. Поэтому количество аскоспор, образуемых на одном плодовом теле, здесь нередко измеряется миллионами (например у крупных пециц или сморчков).
Таким образом, открытое положение гимения наиболее обеспечивает максимальную продукцию и активное рассеяние аскоспор по мере их созревания. Однако такое положение представляет вместе с тем некоторые опасности повреждения гимения во время его развития. Оно устраняется отчасти тем, что гимений залагается сначала только из парафиз, а наиболее важные органы его — сумки лишь позднее, по мере созревания, придвигаются снизу, достигая своими концами верхней границы гимения только к моменту почти полной зрелости. Для этих же целей защиты молодых сумок служит также нередко образуемый из концов парафиз эпитеций. Наконец, во многих случаях молодой гимений защищен еще более массивными тканями самого плодового тела в виде специально кроющего слоя (у Phacidiales и Hysteriales) или в виде общей ткани его, когда плодовое тело в молодости замкнуто (у многих Pezizales). Такое гемиангиокарпное развитие гимения, возможно, унаследовано некоторыми группами дискомицетов от их предков, близких к пиреномицетам.
Если теперь обратиться собственно к филогенетическим отношениям дискомицетов, то нужно прежде всего отметить, что у них намечаются довольно близкие и, повидимому, родственные связи с пиреномицетами. Даже самая граница между этими разделами довольно условна и определяется различно разными авторами. Например, Гейман (1926) помещает Hysteriales среди пиреномицетов, a Hemisphaeriales среди дискомицетов, а у нас имеется как раз обратное размещение этих порядков. Во всяком случае это указывает на неясность границы.
Hysteriales, повидимому, и филогенетически связываются с пиреномицетами, в частности с Sphaeriales, с которыми они сближаются по наличию темного пигмента, а также некоторыми переходными формами, как упомянутые Lophiostomaceae с их щелевидным отверстием перитеция (см. стр. Порядок Hysteriales). Таким образом, апотеций Hysteriales, повидимому, филогенетически выводится из перитеция. Однако нам кажется более правильным все-таки помещать этот порядок среди дискомицетов, так как он представляет дальнейший этап эволюции сумчатых грибов, выразившийся в развитии гимения.
Аналогичный ход эволюции замечается также в порядке Phacidiales. Повидимому они связываются филогенетически с Hemisphaeriales. Строение перитеция некоторых представителей последних (например, Microthyrium) с радиально построенным кроющим слоем довольно сходно с тем, что считается характерным для Phacidiales, кроющий слой которых разрывается радиальными трещинами. С другой стороны, имеются переходные формы между Hysteriales с их вытянутыми апотециями и Phacidiales, апотеции которых имеют в общем округлое радиальное строение. Повидимому, между этими двумя порядками также имеются родственные отношения. Имеются они, как кажется, и между Pezizales и Phacidiales. Поэтому вероятно, что порядок Phacidiales должен рассматриваться как филогенетически сборный.
Что касается, наконец, Pezizales, то положение этого важнейшего порядка дискомицетов весьма неясно. По сложению плодовых тел, имеющих в основном мясистую консистенцию, они напоминают IJypocEeales. Однако каких-либо родственных связей с ними здесь не замечается. Возможно, что здесь имеется только совпадение. С другой стороны, среди низших сумчатых грибов известен упомянутый выше (стр. Порядок Exoascales) Ascocorticium. По общему сложению слабо развитого плодового тела со слоем сумок на его поверхности он напоминает простейших Pezizales (сем. Pyronemaceae). К сожалению, Ascocorticium совершенно не изучен со стороны истории развития и цитологии. Поэтому нельзя судить о том, представляет ли он примитивную форму или редуцированную, произошедшую из высших дискомицетов. В первом случае он мог бы, может быть, рассматриваться как исходная форма для эволюции Pezizales. Однако нужно указать, что промежуточных форм в этом направлении неизвестно.
Если так неясно положение порядка Pezizales в целом, то взаимоотношения семейств, входящих в его состав, кажутся яснее. Две группы их, Operculatae и Inoperculatae, отличаются друг от друга не только по способу открывания сумки (крышечкой у первых и разрывом на вершине у вторых), но и целым рядом других свойств. Так, у Operculatae мало развито бесполое размножение, почти совершенно отсутствуют паразитные формы, плодовые тела имеют довольно простое анатомическое строение, гимений не прикрыт особым эпитецием, образованным из концов парафиз. Inoperculatae отличаются противоположными этому признаками. У них обильно представлено бесполое размножение, много паразитных форм, апотеций у некоторых более сложного анатомического строения с особым толстостенным перидием (у Mollisiасеае), у многих имеется хорошо выраженный эпитеций. Нужно думать, что обе указанные группы представляют самостоятельные эволюционные ряды, показывающие вместе с тем значительный параллелизм в развитии. Высшие представители их, как Helvellaceae среди Operculatae и Geoglossaceae среди Inoperculatae довольно сходны по строению плодовых тел, показывающих диференцировку на стерильную ножку и плодущую шапочку. Однако это сходство, повидимому, не есть результат непосредственной близости указанных двух семейств, а указывает только на одинаковый уровень достигнутой ими эволюции в двух разных рядах. Наиболее примитивным семейством Operculatae нужно считать Pyronemaceae, с их с самого начала открытым гимением и слабо развитым гипотецием. В этом последнем отношении зачаточное плодовое тело Ascodesmis напоминает даже Gymnoascaceae (см. стр. Порядок Pezizales). Вместе с этой общей примитивностью Руrоnеmaceae наиболее сохранили первоначальный половой процесс и типичное строение половых органов.
От Pyronemaceae выводятся Ascobolaceae, Pezizaceae и, может быть, Rhizinaceae. Во всех этих семействах плодовые тела более развиты, гимений залагается под прикрытием кроющего слоя и половой процесс показывает различные степени редукции, заменяясь у очень многих представителей различными формами апогамии (см. стр. Половое воспроизведение, смена генераций и ядерных фаз у аскомицетов). Наиболее развито среди них семейство Pezizaceae, которое можно рассматривать как прямой прогрессивный ряд, идущий от Pyronemaceae. Наконец, повидимому, из Rhiziriaceae дальше выводятся Helvellaceae. По расчленению плодового тела последние стоят выше всех других Operculatae и вместе с тем, повидимому, дальше других зашли в своей апогамии.
Аналогичную эволюцию можно подметить и у Inoperculatae, начиная от Patellariaceae и до сморчковидных Geoglossaceae. Однако в подробностях она различно толкуется разными авторами.
Наконец, Tuberales представляют последний из порядков, примыкающих к дискомицетам. Филогенетически они довольно ясно выводятся из Pezizales. Особенности строения зрелых плодовых тел Tuberales носят ясно приспособительный характер и объясняются подземным развитием их.
Изложенные соображения о филогенезе дискомицетов могут быть иллюстрированы нижеследующей схемой.
Необходимо признать, что принятая здесь система дискомицетов, так же как и пиреномицетов, может возбуждать в ряде моментов сомнение относительно ее филогенетической значимости. Поэтому многие новейшие микологи-систематики (Theissen, Petrak, Arnaud, Nannfeldt и др.) делают попытки в корне пересмотреть систему сумчатых грибов. Однако предлагаемые ими новые системы не разработаны в подробностях и не совсем согласны друг с другом. Как пример их можно указать на систему Nannfeldt (1932). Он делит прежде всего плодосумчатые грибы на три эволюционных ряда: Plectascales, Ascoloculares и Ascohymeniales. Первые в общем соответствуют по своему объему обычным представлениям об этой группе. Ascoloculares охватывают такие группы, как Myriangiales, Dothideales, Mycosphaerellaceae, Hemisphaeriales и некоторые другие. Наконец, Ascohymeniales охватывают большинство дискомицетов и пиреномицетов.
Наиболее оригинальна в этой системе группа Ascoloculares. Центральное место в ней для филогенетических построений автора занимают Myriangiales. Он выводит из их loculi плодовые тела и других относящихся сюда групп, рассматривая их оболочки как остатки сохранившейся стромы Myriangiales. Такие взгляды вряд ли можно считать хорошо обоснованными. Прежде всего для этого отсутствуют данные по истории развития Myriangiales, а между тем вопрос о возможных гомологиях их loculi не может быть решен без знания их происхождения (происходят ли они каждый из особого архикарпа или все из одного). Далее, довольно произвольным кажется толкование оболочек отдельных перитециев как остатков стромы. Также и объединение большинства дискомицетов и пиреномицетов хотя и имеет, может быть, некоторые филогенетические основания, но практически вряд ли удобно.
Поэтому, сознавая известную условность старой систематики аскомицетов, мы все-таки предпочитаем здесь придерживаться в основном ее, так как она более разработана и на ней построены большие систематические сводки, которыми каждому приходится пользоваться для справок и т. п.
Порядок Tubekales, или трюфелевых, содержит около сотни видов с подземными плодовыми телами, во взрослом состоянии замкнутыми в виде мясистых, иногда довольно крупных клубней (до 1 кг весом). При всем несходстве этих образований в готовом состоянии с апотециями типичных дискомицетов, история развития показывает значительные черты близости, как это удалось установить в сравнительно недавнее время работами главным образом Е. Fischer и Бухгольца.
Зрелое плодовое тело трюфеля одето бурой многослойной оболочкой — перидием, а находящаяся под ней внутренняя мясистая ткань имеет как бы мраморный рисунок из чередующихся более светлых и более темных прослоек. Первые получили название внутренних вен (venae internae), а вторыe — наружных вен (venae externae). У некоторых видов (Tuber rufum, Tuber excavatum и др.) наружные вены сходятся все к одному пункту — к поверхности плодового тела, где общая оболочка его прерывается, и открываются таким образом наружу (рис. 219, 1). У других видов (Tuber aestivum и др.) таких пунктов имеется на поверхности несколько. Чередующиеся с наружными венами более плотные внутренние вены отходят от наружных частей под общей оболочкой и загибаются в общем к тому же пункту (или пунктам), где сходятся наружные вены, но кончаются свободно.
Рис. 219. Tuber rufum: l — разрез плодового тела; а — перидий, d — наружные вены, с — внутренние вены, h — гимений; 2 — то же при большем увеличении, значение букв такое же.
Очень молодые стадии развития, например у Tuber excavatum, показывают, что плодовое тело залагается в виде блюдцевидного образования, на вогнутой и несколько складчатой поверхности которого появляются зачатки гимения в виде поверхностного слоя палисадно расположенных гиф — парафиз (рис. 220, 2).
Рис. 220. 1 — разрез молодого плодового тела Pachyploeus; 2 — то же Tuber excavatum; 3 — то же Gonea.
В дальнейшем этот зачаток, находящийся в земле, не может свободно разрастаться в ширину, а при своем росте дает дальнейшие складки на гимениальной поверхности и смыкает вместе с тем первоначально широко открытое свое устье до узкого отверстия — того пункта, в котором наружные вены открываются наружу. При этом развитии первоначальные складки развиваются в внутренние вены. Покрывающий их гимений сложен первоначально из одних парафиз, которые сильно разрастаются, переплетаются с такими же парафизами, отходящими от противоположной складки, и заполняют промежутки между ними более рыхлой тканью, образуя таким образом наружные вены. На границе между ними и внутренними венами, между основаниями парафиз, развиваются сумки, которые не образуют здесь особенно правильного слоя, имеют более или менее овальные, а не цилиндрические очертания и содержат часто меньше 8 спор вследствие ранней дегенерации некоторых из них (рис. 219, 2). У Choiromyces при том же первоначальном развитии гимений образуется не на всей поверхности первоначальных складок, а только на более глубоких (основных) участках их, верхние же (дистальные) участки их в дальнейшем вполне срастаются друг с другом, так что venae externae здесь облитерируются. Благодаря этому в зрелом плодовом теле гимений с сумками выстилает замкнутые, не сообщающиеся друг с другом щелевидные полости. С другой стороны, в роде Pachyploeus более сохраняются первоначальные черты строения и на взрослом плодовом теле. Здесь оно сохраняет довольно широкое устье на вершине, в которое открываются все наружные вены, заполненные, как и у Tuber, рыхлой плектенхимой (рис. 220, 1).
Во всех разобранных примерах заложение гимения происходит по гимнокардному типу, и они выводятся эволюционно из таких форм, как Rhizinaceae. Несколько иначе происходит развитие у Genea и некоторых других. Здесь гимений залагается гемиангиокарпно под прикрытием сплетения гиф. Затем плодовое тело, разрастаясь, делается блюдцевидным и чашевидным, и несущая гимений поверхность делается вогнутой (рис. 220, 3, a, b). Позднее плодовое тело совсем замыкается в полый шар с небольшим отверстием, и внутренняя поверхность его стенки несет гимений, прикрытый со своей морфологически верхней стороны (здесь обращенной к центру плодового тела) первоначальным кроющим сплетением и эпитецием, образованным из верхних концов парафиз. Самый гимений здесь наиболее сохраняет свое дискомицетное строение с цилиндрическими сумками и аскоспорами в них, расположенными в один ряд. У некоторых видов Genea несущая гимений поверхность дает складки, так что получается в готовом состоянии не полое плодовое тело, а отчасти выполненное этими складками. Genea и близкие к ней могут быть выведены из Pezizaceae.
Tuberales представляет прекрасный пример влияния условий существования на строение плодового тела. Имеются значительные черты сходства между ними, Terfeziaceae и Elaphomycetaceae (см. стр. Порядок Plectascales (Плектасковых)). Однако это сходство представляет конвергенцию — результат одинаковых условий (подземные плодовые тела). Плодовые тела трюфелевых вторично замыкаются, и сумки развиваются внутри в защищенные от давления почвы общим перидием. Типичный гимений дискомицетов утрачивает у них свои первоначальные черты: сумки теряют свою цилиндрическую форму и правильное расположение. Все это объясняется тем, что гимению здесь не приходится рассеивать свои споры, которые освобождаются при общем разрушении плодового тела под землей или при поедании его животными (особенно роющими грызунами).
Распространение Tuberales
По распространению Tufcerales связаны с древесными породами, где являются, повидимому, обязательными микоризообразователями. В связи с этим споры их прорастают с чрезвычайным трудом, и культура их не удается. Во всяком случае плодовых тел в культуре до сих пор не получено. Это создает большие трудности изучения их истории развития, так как приходится выискивать ранние стадии в естественных местообитаниях между частицами почвы, что представляет большие затруднения. Вследствие этого, хотя трюфелевые издавна привлекали внимание микологов, история развития их изучена далеко не полно. Между прочимг здесь мало данных по самым ранним стадиям и цитологии.
Практическое значение Tuberales
Важнейшими представителями Tuberales являются виды Tuber, имеющие высокую ценность как съедобные грибы. Из них особенно важен черный, или французский, трюфель (Tuber brumale var. melanosporum). Он встречается особенно во Франции в дубовых лесах на известковой почве и имеет большое промышленное значение. Ценится также, хотя и менее, Tuber aestivum, который встречается и у нас на юго-западе и кое-где в Закавказьи. Под Москвой, особенно в окрестностях Загорска, довольно распространен белый трюфель (Choirоптуces meandriformis). Его крупные, похожие на картофель, плодовые тела при созревании слегка выступают из земли. Съедобен, но не имеет такой ценности, как Tuber.
К Pezizales относятся более 3000 видов как паразитов, так и главным образом сапрофитов.
Эта наиболее типичная группа среди дискомицетов характеризуется главным образом тем, что плодовые тела здесь мясистые, в большинстве случаев блюдцевидной формы, нередко ярко окрашенные. На верхней, обыкновенно более или менее вогнутой, поверхности они несут гимений. По развитию они чаще всего гемиангиокарпны и представляют в молодости полузамкнутые полые тела с узким отверстием на вершине (и даже совсем замкнутые, без отверстия). В этом состоянии они напоминают строение перитеция, отличаясь, однако, тем, что сумки расположены внутри них широким вогнутым слоем — гимением, а не узким пучком у основания плодового тела, как это типично для перитециев. Позднее при созревании эти замкнутые плодовые тела разворачиваются в блюдцевидное тело с открытым гимением. Реже гимений располагается на выпуклой поверхности плодового тела (например у Geoglossaceae и Helvellaсеае). В этом последнем случае они и по развитию оказываются иногда вполне гимнокарпными, т. е. гимений залагается поверхностно ничем не прикрытым (например у Geoglossum) или залагается гемиангиокарпно под прикрытием кроющего сплетения гиф, облекающих весь зачаток плодового тела и при созревании ослизняющихся и исчезающих (Helvella, Spathularia). Плодовые тела обычно образуются непосредственно на мицелии или вырастают из склероциев. Значительно реже встречаются здесь особые стромы, на которых развиваются апотеции (например у Bulgaria, Cyttaria и некоторых других). Кроме сумчатых спороношений, у некоторых Pezizales имеются конидиальные, но последние далеко не достигают такого развития, как это свойственно, например, многим пиреномицетам.
Со стороны ранних стадий развития плодовых тел и половой функции изучено довольно значительное количество Pezizales, особенно представителей некоторых семейств, как Ascobolaceae, Pezizaceae и Pyronemaceae, тогда как большинство других с этой стороны затронуто очень мало. Относящаяся сюда Pyronema confluens представляет классически изученный пример полового воспроизведения у аскомицетов (см. выше, стр. Половое воспроизведение, смена генераций и ядерных фаз у аскомицетов). Довольно сходны с ней в этом отношении Ascodesmis nigricans, Ascobolus vinosus и некоторые другие (рис. 210).
Рис. 210. Ascodesmis nigricans, апотеции.
Здесь также имеется женский половой аппарат из двух многоядерных клеток: аскогона и трихогины, и одноклетный многоядерный антеридий. Оплодотворение и дальнейшее развитие вполне сходно с Pyronema. Этот тип, свойственный простейшим представителям Pezizales, нужно счесть первоначальным для них. В дальнейшей эволюции наблюдается ряд изменений в половой функции и сложении половых органов. У Lachnea stercorea общие очертания архикарпа сходны с Pyronema за исключением того, что трихогина здесь многоклетная. У многих видов Ascobolus, Ascophanus и др. архикарп представлен многоклетной, часто спирально закрученной гифой с многоклетной трихогиной и мало выделяющейся аскогенной клеткой (или несколькими аскогенными клетками, лежащими в виде слабо диференцированной толстоватой гифы), из ряда клеток которой развиваются аскогенные гифы (см. рис. 38, 7). В этих случаях он вполне подходит под обозначение «воронинская гифа».
Рис. 38. Половые органы аскомицетов: 1 и 2 — Ascobolus carbonarius; 2 — начало развития аскогенных гиф из двух клеток аскогона; 3 — Ascobolus magnificus; 4 — PeniciIlium; 5 — Sphaerotheca; 6 — Polystigma; 7 — Rhizina undulata, развитие аскогенных гиф из ряда клеток архикарпа; m — трихогина; к — конидия; ♀ — архикарп; ♂ — антеридий.
Одновременно с этим наблюдается подавление половой функции. У Lachnea stercorea развивается еще нормальный антеридий, и происходит копуляция его с трихогиной; однако дальше из нее в аскогон мужские ядра не проходят. У весьма многих, изученных в этом отношении видов (многие Ascobolus, Humaria и др.), антеридии совсем не развиваются, и дальнейшее развитие архикарпа происходит без цитогамии, апогамически. При этом или происходит переход ядер из одних клеток архикарпа в соседние аскогенные, или первичные дикарионы формируются в пределах одной многоядерной клетки аскогона. В дальнейшем из аскогонов, как полагается, вырастают аскогенные гифы, и на них образуются аски, где и происходит кариогамия двух ядер дикариона. У некоторых представителей, как Humaria granulata, параллельно выпадению цитогамии (и отсутствию антеридия) женский половой аппарат теряет свою трихогину, превращаясь в многоядерную круглую клетку на конце архикарпа. В ней также без цитогамии происходит формирование первичных дикарионов. Наконец, у некоторых Pezizales в связи с потерей половой функции (цитогамии) наблюдается выпадение не только антеридия, но и архикарпа. Это имеет место например у Helvella crispa, Humaria rutilans, Peziza vesiculosa и некоторых других. Здесь аскогенные гифы берут начало из обычных вегетативных клеток плодового тела, в которых благодаря переползанию ядер из соседних клеток возникают первичные дикарионы.
Вообще Pezizales представляют один из наилучших примеров, где можно проследить постепенную утрату половой функции и редукцию половых органов. Правда, здесь, как и в других аналогичных случаях, нередко можно наблюдать у близких форм различные степени этого редукционного процесса, так что очевидно он мог возникать самостоятельно в отдельных мелких группах и поэтому им нельзя пользоваться для установления родственных отношений между последними. Однако в общем надо отметить, что простейшие по всем признакам среди Pezizales, такие как Pyronema, Ascodesmis (сем. Pyronemaсеае), имеют и наиболее типично развитой и функционирующий половой аппарат. В промежуточных семействах, как Ascobolaceae, Pezizaceae, замечаются наряду с типично половыми формами и такие, которые более или менее далеко зашли по пути апогамии (например разные виды родов Ascobolus, Lachnea и др.). Наконец, наиболее специализированные семейства Helvellaceae и Geoglossaceae показывают в общем и наиболее далеко зашедшую редукцию половой функции и половых органов.
Классификация Pezizales
Систематическое подразделение обширного порядка Pezizales производится на основании различных признаков: общего сложения плодового тела, его анатомического строения, гимнокарпности или гемиангиокарпности и, наконец, способа открывания сумок: крышечкой или простым отверстием на вершине. Согласно этому последнему признаку, все Pezizales могут быть разбиты на два ряда. Operculatae (сумки открываются крышечкой) и Inoperculatae (сумки открываются простым отверстием).
В обоих этих рядах можно установить иногда известный параллелизм по другим признакам (например между Geoglossaceae и Helvellaceae по общему сложению и развитию плодового тела).
Таким образом, здесь мы можем указать на следующие важнейшие семейства.
Operculatae
I. Апотеций блюдцевидный с вогнутой гимениальной поверхностью. Эпитеций не образуется. A. Апотеций с самого начала открытый (гимнокарпный). Гипотеций слабо развитой. Перидий не диференцирован. 1. Сем. Pyronemaceae. B. Апотеций с начала большей частью закрытый (гемиангиокарпный). Гипотеций и перидий мало диференцированы друг от друга. 1. Сумки при созревании высовываются над общим уровнем гимения. 2. Сем. Ascobolaceae. 2. Сумки при созревании не высовываются над общим уровнем гимения. 3. Сем. Pezizaceae. II. Апотеций с выпуклой гимениальной поверхностью. A. Апотеций без ножки. 4. Сем. Rhizinaceae. B. Апотеций с ножкой. Гимений только на верхней его части, расширенной в виде шапочки, большей частью с складчатой (морщинистой) поверхностью. 5. Сем. Helvellaceae.
Inoperculatae
I. Апотеций блюдцевидный с вогнутой или плоской гимениальной поверхностью. A. Свободные концы парафиз образуют эпитеций. 1. Апотеций блюдцевидный открытый. 6. Сем. Patellariaceae. 2. Апотеций в молодости полузакрытый, часто на стромах кожистой или слизистой консистенции. 7. Сем. Cenangiaceae. 3. Апотеций в большом числе на мясистых шаровидных стромах. 8. Сем. Cyttariaceae. B. Эпитеция не развивается. 1. Перидий образован из вытянутых, большей частью тонкостенных и светлоокрашенных клеток. 9. Сем. Helotiaceae. 2. Перидий из паренхиматических, чаще всего толстостенных темноокрашенных клеток. 10. Сем. Mollisiaceae. II. Апотеций булавовидный; гимений на выпуклой поверхности в его верхней части. 11. Сем. Geoglossaceae.
1. Сем. Pyronemaceae
Небольшое семейство (около 25 видов), куда относится прежде всего Pyronema confluens — известная как классический пример нормальным половым образом развивающегося аскомицета (рис. 211).
Рис. 211. Pyronema confluens, схема разреза через плодовое тело. Диплоидная фаза изображена жирными контурами, гаплоидная — тонкими контурами.
Этот сапрофитный гриб встречается в природе на почве на месте бывших костров, где его мелкие (около 1 мм в диаметре) красноватые апотеции развиваются обыкновенно группами и поэтому хорошо заметны. Такое местообитание Pyronema обусловлено тем, что в нормальной почве она вытесняется находящимися там почвенными бактериями, попадая же на прогретую и отчасти стерилизованную почву, ее споры прорастают и развивают мицелий с апотециями на нем (стр. Совместная культура с другими микроорганизмами). По тем же причинам Pyronema нередко разводится на поверхности стерилизованной почвы в сосудах для вегетационных опытов.
Ascodesmis nigricans также довольно хорошо изучен со стороны истории развития. Апотеции у него видны только в микроскоп. Они состоят почти из одного только пучка сумок с парафизами и имеют мало развитую окружающую ткань (рис. 210). По общему сложению апотеция Ascodesmis напоминает отчасти Gymnoascaceae. Встречается на навозе, особенно коровьем.
Рис. 210. Ascodesmis nigricans, апотеции.
2. Сем. Ascobolaceae
Типичное копрофильное семейство. Представители его весьма обычны на навозе различных травоядных животных. Плодовые тела обыкновенно очень мелкие, но обладают большей частью очень активно действующими аппаратами, разбрасывающими аскоспоры. В связи с этим у некоторых, как у Ascobolus immersus, аскоспоры отличаются чрезвычайной величиной, а у других (Saccobolus и др.) все восемь аскоспор склеиваются в один пучок, вылетающий из сумки. Из отдельных представителей следует специально отметить Ascobolus magnificus, A. carbonarius и A. stercorarius — интересные своим гетероталлизмом (Dodge, 1920; Betts, 1926; Green, 1931).
Thelebolus stercoreus образует в плодовом теле только одну очень крупную сумку, содержащую до 1000 аскоспор. Аскогенных гиф здесь совсем не развивается, а сумка прямо образуется из одной клетки архикарпа, содержащей два клеточных ядра. Thelebolus интересен еще исторически тем, что в свое время его крупные многоспоровые сумки рассматривались Брефельдом как переход от сумки к спорангию и самый гриб помещался в особую переходную группу Hemiasci. Сейчас вполне выяснено, что аски Thelebolus развиваются типично с кариогамией и последующим редукционным делением ядра. Их многоспоровость есть результат увеличения числа ядерных делений в сумке (с 3 до 10).
Ascobolaceae легко получаются в культуре и дают здесь плодовые тела. Поэтому они много исследовались со стороны истории развития и цитологии, так что из 150 приблизительно известных видов изучено с этой стороны не менее 15. Такого высокого процента пожалуй нет ни в какой другой группе грибов.
3. Сем. Pezizaceae
Около 700—800 видов главным образом сапрофитов на: почве, древесине, некоторые на навозе травоядных животных. Плодовые тела мясистые, у некоторых довольно крупные (несколько сантиметров в диаметре). Главные роды Lachnea, Otidea, Peziza (рис. 212). Последний, насчитывающий больше 400 видов, делится на несколько подродов: Humaria, Pustularia, Galactinia, Macropodia, Acetabula и др. Два последние отличаются тем, что плодовые тела у них приподняты на центральной ножке, что представляет отчасти переход к семейству Helvellaceae.
Рис. 212. 1 — Lachnea hemisphaerica — разные стадии развития апотеция; 2 — Humaria rutilans; 3 — Macropodia (все в естественную величину).
Из отдельных представителей особый интерес представляет Humaria granulata. Этот, нередко встречающийся, копрофильный гриб по исследованиям Gwynne Vaughan и Williamson (1930) оказался гетероталличным. Однако гетероталлизм здесь представляет ту особенность, что из двух имеющихся сортов аскоспор те и другие дают мицелии с архикарпами (оогониями) и ни одна не дает антеридиев. Однако в односпоровой культуре архикарпы замирают, и, плодовых тел не развивается. Развитие происходит лишь при соединении двух различных мицелиев. Тогда на месте соединения их образуются многочисленные анастомозы, и залагающиеся на том и другом архикарпы развиваются дальше апогамически и дают апотеции. Таким образом, гетероталлизм Humaria granulata не может быть представлен просто как раздельнополость. Можно представить эти своеобразные отношения таким образом, что по существу все мицелии обоеполые, точнее, содержат обе половые потенции, и коих, однако, внешне выявляется только женская. Противоположные мицелии действуют друг на друга своимискрытыми мужскими зачатками, которые бессильны по отношению к собственным оогониям, но возбуждают развитие оогониев «другого знака». Возможно, что это в конце концов сводится к действию гормонов, различных в мицелиях (+) и (—), аналогично тому, что указывается для Neurospora (см. стр. Гипотеза Гартмана).
Некоторые Pezizaceae дают в культуре довольно обильные конидиальные спороношения. Таковы: Peziza vesiculosa, с неветвящимися, вздутыми на концах, конидиеносцами типа Oedocephalum или Lachnea cretea также с вздутыми на концах, но ветвящимися конидиеносцами.
4. Сем. Rhizinaceae
Небольшое семейство (около 20 видов). Главный представитель его, Rhizina inflata, довольно обычен в лесах на почве, где живет, повидимому, чаще сапрофитно, но иногда и паразитно на древесных корнях. Плодовое тело этого гриба плоское, слегка выпуклое сверху, где расположен гимений. На ранних стадиях развития плодового тела в нем замечается несколько мало диференцированных архикарпов в виде воронинских гиф. Сюда же относится Sphaerosoma. В молодости ее плодовое тело блюдцевидно, с гимением на верхней стороне. Потом гимениальная сторона сильно разрастается, и плодовое тело принимает приблизительно шаровидный вид с гимением на поверхности.
5. Сем. Helvellaceae
Сюда относится около 100 видов сапрофитных грибов, растущих на почве. Плодовые тела — большей частью крупные, диференцированные на пенек и шляпку. Только эта последняя несет на своей наружной, выпуклой и часто морщинистой поверхности гимений. Последний залагается, поскольку это выяснено у ряда форм, гемиангиокарпно под прикрытием общей оболочки, позднее исчезающей. Важнейшие роды, Helvella, Gyromitra, Verpa, Morchella (рис. 213) — довольно важные съедобные грибы (сморчки, строчки). Поверхность шляпки у трех последних складчатая, морщинистая. Особенно глубоко и правильно идут эти складки у Morchella, разбивая поверхность шляпки на ряд глубоких открытых снаружи камер. У Morchella только они выстланы гимением, а отделяющие их ребра (края складок) остаются стерильными, так что общий гимений разделяется на отдельные участки.
Рис. 213. Verpa bohemica.
6. Сем. Patellariaceae
Около 250 видов. Сюда относятся частью сапрофиты (преимущественно на древесине), частью паразиты (особенно на лишайниках). Плодовые тела — мелкие, обычно темно окрашенные, кожистые; у сапрофитов они развиваются чаще всего поверхностно, а у паразитов нередко сначала погружены в субстрат и лишь позднее выдвигаются на поверхность. На гимений — хорошо развитой эпитеций из концов парафиз. У некоторых (например Hysteropatella) плодовые тела вытянутые и раскрываются щелью, напоминая этим Hysteriales. Co стороны истории развития Patellariaceae не изучены. Patellaria — на древесине, Scutula — паразиты на лишайниках.
7. Сем. Cenangiaceae
Сюда относится больше 500 видов, отчасти сапрофитов, преимущественно на мертвых древесных ветвях или иногда на травянистых стеблях, отчасти паразитов на деревьях. Плодовые тела образуются часто на особых стромах, залагающихся внутри субстрата и затем выдвигающихся на поверхность. Строма—слизистой или кожистой консистенции. У многих на той же строме сначала развиваются конидиальные спороношения: пикниды (например у Dermatea cerasi; рис. 214) или свободные ложа (например у Dermatea carpinea).
Апотеции сначала прикрыты кроющим слоем, который затем при созревании исчезает или разрывается лопастями, напоминая этим Phacidiales. Кроме того, имеется хорошо развитой эпитеций, образованный кончиками парафиз. Цитологически Cenangiaceae не изучены. Главнейшие представители: Cenangium abietis — на ветвях хвойных деревьев как сапрофит и иногда как паразит, вызывающий их отмирание. Dermatea carpinea — на ветвях граба, Dermatea cerasi — на видах Prunus. Сюда же можно отнести нередко выделяемые в особое семейство Bulgariaceae роды Bulgaria, Sarcosoma и др. Bulgaria polymorpha — на мертвых ветвях дуба — интересна обилием и разнообразием конидиальных спороношений, а также тем, что в сумках из 8 спор 4 более крупные темно окрашенные и 4 более мелкие светлые (эта разноспоровость имеет своей основой то, что ядерные деления в аске происходят не одновременно, и около 4 позднее возникших ядер формируются более мелкие споры). Близко к Bulgaria стоит Sarcosoma globosum. Ее крупные (до 5 и больше сантиметров в диаметре) шаровидные плодовые тела развиваются прямо на почве. На верхней приплюснутой стороне их расположен диск гимения; нижняя часть одета довольно прочной кожистой пленкой, а внутри образована чрезвычайно водянистой тканью. Этот оригинальный и считающийся редким гриб встречается в больших количествах весной в сосновых лесах в некоторых местах под Москвой (Быково, Влахернская).
8. Сем. Cyttariaceae
Это семейство представлено одним только родом Cyttaria, несколько известных видов которого паразитируют на ветвях буков (Notholagus) в южном полушарии. Здесь очень характерны стромы, развивающиеся группами на ветвях и представляющие мясистые, в большинстве случаев ярко окрашенные шаровидные тела размером с горошину. На их поверхности в особых углублениях развиваются многочисленные апотеции, прикрытые в молодости кроющим сплетением гиф (рис. 215). Стромы Cyttaria съедобны, и еще Дарвин в «Путешествии на Бигле» указывал, что они составляют существенную часть питания туземцев Огненной Земли.
Рис. 215. Cyttaria Gunnii: a — группа стром на ветви бука; b — разрез стромы.
9. Сем. Helotiaceae
Около 1200 видов сапрофитов и паразитов. Плодовые тела залагаются поверхностно, реже погружены в начале в субстрат. Они блюдцевидной или бокаловидной формы, в молодости замкнутые. У некоторых плодовые тела на ножках. Отличительным признаком Helotiaceae от следующего семейства Mollisiaceae является строение перидия (наружных слоев гипотеция); он образован здесь тонкостенными вытянутыми клетками. Цитологически Helotiaceae не изучены.
Важнейший относящийся сюда род Sclerotinia с несколькими десятками паразитных видов. Как показывает название, для Sclerotinia характерно образование склероциев, являющихся здесь обязательными членами цикла развития. Из них после периода покоя вырастают апотеции, снабженные более или менее длинными ножками. Sclerotinia делится на два подрода: Stromatinia, у которой склероции образуются из пронизанных грибными нитями и мумифицированных плодов, и Eusclerotinia, у которой склероции образуются большей частью поверхностно на вегетативных органах или иногда внутри их, но представляют только грибное сплетение без мумифицированных тканей хозяина (рис. 216 и 217).
Поскольку изучено, эти два подрода отличаются и по имеющимся у них конидиальным спороношениям. У Stromatinia они типа Monilia, а у Eusclerotinia — типа Botrytis. В первом случае образуются короткие неветвящиеся конидиеносцы, и на них развиваются длинными базипетальными цепочками крупные овальные конидии. Между ними при созревании появляются веретеновидные диъюнкторы, образованные на счет среднего слоя перегородки между соседними конидиями (рис. 216, 2). В случае Botrytis конидиеносец моноподиально ветвится, и на концах разветвления развивается головка конидий (не цепочками) (рис. 217, 2).
Практическое значение и биологические особенности Sclerotinia
Представители Sclerotinia важны в практическом отношении и интересны по биологическим особенностям. Из подрода Stromatinia особо любопытны виды, паразитирующие на Vacciniaceae и некоторых близких к ним растениях. Например, очень обычная S. vaccinii, живущая на бруснике, весной заражает молодые побеги хозяина и вызывает их отмирание. При этом на них развиваются конидиальные спороношения типа Monilia, и конидии их заражают распускающиеся к тому времени цветы брусники. Мицелий гриба проникает в завязь и мумифицирует ее. Она сваливается и после зимовки развивает апотеции, аскоспоры которых заражают опять молодые побеги хозяина. Таким образом, здесь конидиальиое спороношение и склероции приурочены к разным органам питающего растения в соответствии с порядком их развития в начале лета. У S. ledi (S. heteroica) и S. rhododendri дело пошло дальше в том отношении, что здесь развитие гриба связывается уже с разными растениями. Именно, первый заражает весной листья голубики, а второй — черники. Конидии, развивающиеся на них, заражают затем распускающиеся к тому времени цветы у первого — багульника (Ledum), а у второго — Rhododendron. Таким образом, здесь вырабатывается разнохозяйственность, зачатки которой мы видели также у Glaviceps (см. стр. Базидиомицеты) и которая в более определенной и обязательной «форме представлена у Uredinales (см. ниже, Uredinaies). Из других Stromatinia большое практическое значение имеют S. cinerea — на плодах слив и других косточковых и S. fructigena — на яблонях и грушах. Здесь на плодах образуются и конидиальные спороношения, передающие заражение на соседние плоды, и позднее здесь же образуются зимующие склероции.
Из подрода Eusclerotinia особенно важны: S. libertiana на очень различных растениях (подсолнечник, георгины, различные овощи, особенно при зимнем их хранении и т. д.), S. trifoliorum — на клевере, S. fuckeliana — на виноградной лозе.
Из других Helotiaceae следует упомянуть Dasyscypha Willkommii, паразит на лиственнице, вызывающий у нее так называемый «рак». Далее, большой род Helotium (около 250 видов), виды которого живут сапрофитно на растительных остатках (главным образом на ветвях, пнях и т. п.)
10. Сем. Mollisiaceae
Сюда относятся 500 видов сапрофитов и паразитов. Отличие от сем. Helotiaceae заключается здесь главным образом в строении перидия апотеция, сложенного у Mollisiaceae из паренхиматических толстостенных темно окрашенных клеток. Представителем может служить Pseudopeziza ribisr паразит смородины. На живых листьях он развивает конидиальные спороношения (типа Gloeosporium), а на перезимовавших уже развиваются апотеции. Сходное развитие имеет Fabraea fragariae — распространенный паразит земляники, вызывающий пятнистость ее листьев (в конидиальном состоянии — Marsonia fragariae). Относительно Fabraea ranunculi (на листьях различных лютиковых) имеются и некоторые цитологические данные: при заложении апотециев (здесь на живых листьях) появляются группами многоклетные с длинными трихогинами архикарпы; антеридиев не развивается (Гусева, 1923).
11. Сем. Geoglossaceae
Около 100 видов преимущественно сапрофитов. Плодовые тела булавовидные или головчато утолщенные на конце. Эта расширенная часть покрыта гимением. Последний у одних представителей залагается гимнокарпно (Geoglossum), а у большинства других — гемиангиокарпно под общей обверткой плодового тела, позднее исчезающей. На ранних стадиях развития плодового тела в ряде случаев наблюдались архикарпы, в виде мало диференцированных воронинских гиф и без антеридиев.
Главнейшие представители: Geoglossum — плодовые тела булавовидные (рис. 218, 1); Leotia — плодовые тела на вершине несут головку, резко отграниченную от ножки и покрытую гимением (рис. 218, 2); Spathularia — плодовые тела имеют форму лопаточки, на ней плодущая часть развита с двух противоположных сторон.
Характерным признаком Phacidiales является то, что гимений в плодовом теле долго остается закрытым и открывается наконец вследствие разрыва лопастями прикрывающего его слоя или у некоторых широкой щелью. Сюда относится около 600 видов преимущественно сапрофитов и отчасти паразитов, причем последние образуют свои сумчатые спороношения обычно лишь на отмершей ткани, т. е. сапрофитно.
Как наиболее известные и лучше изученные примеры можно привести следующие. Rhytisma acerinum — паразит на листьях клена, образующий на них черные блестящие пятна (рис. 209, 1).
Рис. 209. 1 — Rhytisma acerinum на листе клена; 2 — Clithris quercina на ветви дуба.
Гифы гриба распространяются здесь преимущественно в клетках верхнего эпидермиса, разрушая его боковые стенки и сплетаясь благодаря этому в сплошное плоское ложе. На его поверхности в средине лета отдельными тесными группами развиваются конидиеносцы и отшнуровывают конидии, прорывающие лежащую над ними верхнюю стенку эпидермиса. Конидии здесь, повидимому, не играют роли в дальнейшем распространении гриба. Во второй половине лета конидиальные спороношения исчезают, а стромы утолщаются и чернеют, приобретая характер склероциев. Осенью в них появляются плоские полости — будущие апотеции, и в каждой из них залагается по нескольку архикарпов, составленных из 2—3 многоядерных клеток. Антеридиев совсем не развивается, и оплодотворение заменяется переходом ядер из 1—2 клеток архикарпа в одну более крупную клетку ее, являющуюся аскогоном. После опадения и зимовки листа эти аскогоны развивают аскогенные гифы и на них сумки, каждая с 8 вытянутыми почти нитевидными одноклетными аскоспорами. При созревании апотециев кроющий темно-окрашенный слой над ними с соответствующим участком верхней стенки эпидермиса разрывается и несколько отгибается к краям лопастями, обнажая светлый гимений. Аскоспоры заражают снова молодые листья клена (Jones, 1925).
Сходное в общем развитие имеет Cryptomyces pteridis, нередко встречающийся у нас на папоротнике орляке. Гифы гриба распространяются здесь по всему листу и местами образуются плоские ложа под эпидермисом против устьиц. На них развиваются тесным слоем конидиеносцы. Образуемые ими конидии могут давать здесь новое заражение орляка. К концу лета на пораженном листе под эпидермисом образуются новые, более массивные, сплетения — зачатки апотециев. В них также диференцируются многоклеточные архикарпы, но не развивается антеридиев. Оплодотворение заменяется, по указаниям Rillian (1918). копуляцией двух соседних архикарпов, причем ядра из одного переходят в другой. После зимовки из этого последнего развиваются аскогенные гифы и аски. Раскрытие апотеции происходит так же, как у Rhytisma. Phacidiales могут быть разделены на 3 семейства.
Сем. Stictidaceae — с мясистыми светлоокрашенными плодовыми, телами. Ocellaria aurea на мертвых ветвях.
Сем. Tryblidaceae — с темными апотециями, развивающимися на выступающей из субстрата строме. Scleroderris ribesia на мертвых ветвях смородины.
Сем. Phacidiaceae — с темными апотециями на плоской, погруженной в субстрат строме. Rhytisma. Cryptomyces. Clithris quercina на мертвых ветвях дуба (рис. 209, 2).
Этот порядок занимает отчасти промежуточное положение между дискомицетами и пиреномицетами. К нему относится около 500 видов как паразитов, так и сапрофитов, преимущественно на растительных субстратах.
Плодовые тела Hysteriales поверхностные или погруженные в субстрат, черного снаружи цвета и обыкновенно имеют вытянутую линейную форму, нередко извилистую и даже разветвленную. Сверху они прикрыты темным плотным сплетением гиф (кроющий слой), под которым и развивается гимекий. Наиболее характерно для всего порядка то, что при созревании сумок кроющий слой разрывается узкою продольной щелью. Ее края расходятся во влажную погоду, обнажая светло окрашенный гимений. В сухую погоду они снова сближаются к закрывают гимений. По расположению сумок широким слоем — гимением, Hysteriales могут быть отнесены к дискомицетам. С другой стороны, позднее открывание гимения, неполное и временное его обнажение после созревания указывают на близость их с пиреномицетами, в частности с Sphaeriales, где в одном небольшом семействе Lophiostomaceae отверстие перитеция сплюснуто щелевидно. Поэтому некоторые авторы (например Gaumann, 1926) помещают Hysteriales среди пиреномицетов.
В качестве примера Hysteriales можно указать Lophodermium, виды которого очень распространены у нас как паразиты на лиственных и особенно хвойных деревьях, вызывая отмирание и отпадение листвы (Schutte немецких лесоводов). Наиболее, важен L. pinastri, паразитирующий на хвое сосны и вызывающий массовую гибель сосновых сеянцев в питомниках (рис.208).
Рис. 208. Lophodermium pinastri: а — плодовые тела на хвое сосны; b — то же, слабо увеличено.
В паразитных условиях он развивает на другой год после заражения конидиальные спороношения (пикниды) и уже на отмерших листьях — сумчатые плодовые тела. Из других видов широко распространены: L. macrosporum — на ели, L. nervisequum — на пихте и др. Из других представителей можно назвать Hypoderma, виды которой часто встречаются как сапрофиты на отмерших стеблях травянистых растений, образуя на них черные вытянутые бляшки Н. commune — на злаках.
Близкие к Hysteriales грибы входят в состав лишайников Graphidaceae. Например, у Graphis scripta сумчатые спороношения имеют вид черных ветвящихся черточек, напоминающих восточные письмена.