Явление раздельнополости у грибов впервые установлено Блексли в работах 1904 и 1906 гг.
Оказалось, что у большинства видов изучавшихся им мукоровых на мицелии, выращенном из одной споры, не образуется зигот. Они развиваются лишь при встрече двух мицелиев, притом не всяких, а только некоторых, хотя и одинаковых по внешности, но отличающихся друг от друга в половом отношении. На месте соприкосновения их на том и на другом залагаются органы оплодотворения, и в результате их слияния образуются зиготы (рис. 41, 1).

1 — Phycomyces образование половых органов на месте встречи (+) и (—) мицелиев;
2 — схема образований гетерокариотического мицелия в опыте Burgeff;
S — схема копуляции гомоталличной Absidia (в середине) с гетероталличным (+) и (—) Mucor (справа и слева).
Так как самое оплодотворение у мукоровых грибов обыкновенно имеет строго изогамный характер, то здесь не приходится говорить о мужском и женском мицелиях; поэтому Блексли обозначил их умадно знаками: одни (+), а другие (—), так же, как и производящие их споры, а самое явление такой раздельнополости назвал гетероталлизмом. В дальнейшем раздельнополость была установлена и у некоторых таких мукоровых, где, как у Absidia и др., имеются довольно ясные различия в величине и строении копулирующих органов (рис. 34, 2).
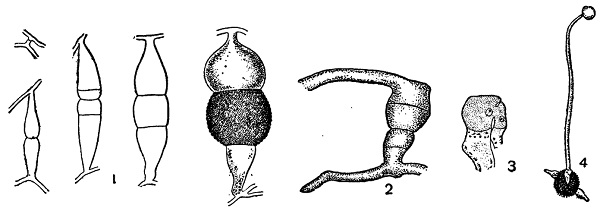
1 — Rhizopus nigricans;
2 — Absidia spinosa — гетерогамная копуляция;
3 — Endogone — гетерогамная копуляция одноядерных клеток;
4 — Мисоr — прорастание зигоспоры образованием зародышевого спорангия.
При этом при встрече соответственных двух мицелиев на одном из них всегда образуются большие (женские), а на другом меньшие (мужские) органы. В этом случае неопределенные обозначения мицелиев (+) и (—) могут быть заменены названиями женский и мужской, а обозначение гетероталлизм обычным — раздельнополость. Однако терминология Блексли столь укоренилась, что употребляется и сейчас почти постоянно для обозначения явлений распределения пола у низших растений (не только у грибов, но и у водорослей). Таким образом, гетероталлизм = раздельнополость, противоположное понятие гомоталлизм = обоеполость.
Гетероталлизм установлен среди всех основных групп грибов. Большинство оомицетов, повидимому, гомоталличны. Гетероталлизм среди них установлен только у немногих, например у нескольких видов Dictyuchus (D. sterilis) и некоторых Phytophthora, как Ph. Faberi и Ph. omnivora, причем у последней, наряду с гетероталличными, обнаружены также и гомоталличные особи. Наоборот, среди зигомицетов гетероталлизм, повидимому, преобладает. По крайней мере среди мукоровых, особенно подробно изученных в указанном отношении, около 60 видов оказалось гетероталличными и около 25 — гомоталличными.
Среди огромной группы сумчатых грибов гетероталлизм установлен до сих лор приблизительно у 25 видов, из коих половина приходится на представителей своеобразных Laboulbeniales. Из других групп сумчатых грибов гетероталличны: Taphrina epiphylla, Taphrina Klebhnü, Penicillium luteum, Ceratostomella coerulea, Neurospora sitophila, Ascobolus magnificus, Ascobolus carbonarius, Ascobolus stercorarius, Humaria granulata и некоторые другие. Нужно, однако, помнить, что аскомицеты только что начали изучать в этом отношении и, вероятно, в дальнейшем будет обнаружено немало еще гетероталличных форм среди них.
При культуре аскомицетов нередко приходится сталкиваться с таким явлением, что гриб в чистой культуре, несмотря на наилучшие как будто условия, упорно не дает сумчатых стадий, тогда как в естественной обстановке в природе они обычны у данной формы. Весьма вероятно, что в ряде случаев это различие имеет в своей основе гетероталлизм: в природе, где имеется разнородный споровой материал, легко может произойти встреча двух подходящих мицелиев, необходимых для развития сумчатых плодоношений, в чистой же культуре, выводимой, как правило, из одной споры, такие условия создаться не могут. Возможно, далее, что гетероталлизм лежит в основе некоторых Fungi imperfecti. В пределах этой огромной группы (около 25 тысяч видов) для многих видов установлено, что они являются только бесполыми стадиями тех или иных аскомицетов. Самая редкость сумчатых спороиошеиий в ряде случаев может быть объяснена их гетероталлизмом и тем, что разные полы имеют неодинаковое распространение. Наконец, возможно, что у некоторых форм один пол совершенно утрачен (или сделался крайне редким); тогда получается настоящий несовершенный гриб.
У базидиомицетов гетероталлизм особенно распространен. Среди головневых изучено в этом отношении больше пятнадцати видов, и все они оказались гетероталличными; весьма вероятно, что таковыми окажутся, если не все, то огромное большинство головневых.
У ржавчинных гетероталлизм установлен впервые только в 1927 г. (Craigie), но с тех пор изучено в этом отношении около 10 видов и 9 из них оказались гетероталличными. Также в самое последнее время обнаружен гетероталлизм у некоторых аврикуляриевых и дрожалковых (Auricularia Auricula Judae, некоторые Exidia).
Среди гименомицетов из изученных форм более пятидесяти оказались гетероталличными и только девять — гомоталличными. Вероятно, гетероталлизм здесь преобладает. Наконец, и у гастеромицетов, когда стали овладевать культурой и этих грибов, в последнее время также обнаружен гетероталлизм (у Sphaerobolus, Lorenz, 1933; Cyathus и Crucibulum, Fries, 1936).
Вообще можно сказать, что гетероталлизм для гаплоидной стадии, к которой, собственно, и относится это понятие, кажется более естественным и понятным, если считать, что определение половых свойств происходит генотипически при редукционном делении. В тех же случаях, где мы имеем явление гомоталлизма, или обоеполости, она может иметь своей основой или фенотипическое определение половой функции, не связанное с генами, локализирующимися в ядре (истинная обоеполость), или в основе ее может лежать так называемая гетерокариотичносць мицелия, т. е. содержание в нем одновременно как (+), так и (—) ядер. Первый тип, повидимому, более распространен. Он устанавливается, например, у большинства гомоталличных мукоровых: Sporodinia, Zygorhynchus, некоторых видов Mucor, Absidia. Это доказывается, между прочим, явлениями регенерации половых органов (до их слияния), что можно вызвать легким повреждением их. При этом каждый из них регенирует в мицелий, и на нем снова образуются копулирующие друг с другом половые органы. Таким образом доказывается, что даже ядра половых органов до их слияния не имеют окончательно фиксированной диференцировки в половом отношении. Что же касается второго типа гомоталлизма (гетерокариотичного), то таковой с достоверностью установлен у Phycomyces, где он получен экспериментально переливанием протоплазмы (с ядрами) из ( —) мицелия в (+) мицелий (Burgeff, 1915, рис. 41, 2). Такой мицелий обычно совсем не дает половых органов и не реагирует ни с (+), ни с (—), но иногда (очень редко) на нем одном образуются зиготы, вероятно, вследствие разделения или отмучивания разнородных ядер в один и другой из копулирующих органов. В естественной обстановке такие гетерокариотические мицелии должны возникать из первично многоядерной споры, содержащей и (+) и (—) ядра, или в результате анастомозов двух мицелиев. Судя по всему, такую же гетерокариотическую основу имеет гомоталличность Neurospora tetrasperma, где в отличие от близких N. sitophila и N. crassa (гетероталличных) аскоспоры залагаются как двуядерные образования (не сестринские ядра) (Dodge, 1927—1928; Wilcox, 1928) (рис. 40).

1 — аскоспора с дикарионом, ядра разных половых знаков, даны светлым и черным;
2 — гетерокариотический мицелий, клетки с одним или несколькими дикарионами; кое-где на конце дикарионы разъединяются и получаются одноядерные клетки.
Также у Pleurage anserina, гдe в сумке нормально залагается по 4 двуядерные споры, они вырастают в гомоталличный мицелий. Но иногда у этого вида вместо одной двуядерной споры развиваются две карликовые одноядерные. Эти последние уже прорастают одна в мицелий (+), другая в мицелий (—). Отсюда можно заключить, что гомоталлизм Pleurage anserina, берущий свое начало от двуядерных аскоспор, имеет в своей основе гетерокариотическое сложение мицелия (Dowding, 1931). Реакция, наблюдаемая при соединении двух разнородных мицелиев гетероталличных видов в случае гименомицетов, которые совершенно лишены половых органов, заключается в появлении анастомозов между их ветвями и переходе ядер из одной анастомозирующей клетки в другую. Таким образом создаются первые двуядерные клетки, которые, разрастаясь, и дают начало двуядерному мицелию. В других же случаях, где имеется настоящий половой процесс (например у мукоровых или Dictyuchus), реакция выражается в появлении на мицелиях половых органов, которые затем и сливаются друг с другом (рис. 41, 1). Любопытно, что такое образование половых органов происходит раньше, чем два мицелия придут в непосредственное соприкосновение. Таким образом, здесь имеется известное действие на расстоянии. У мукоровых это действие передается даже через пленку коллодия, отгораживающую два разнородных мицелия, и через воздух. Отсюда следует, что здесь имеет место возбуждающее действие веществ, с одной стороны, растворимых и диффундирующих через коллодий, а с другой — летучих (Burgeff, 1924).
Мицелии (+) и (—) реагируют друг с другом не только в том случае, когда они принадлежат к одному виду гриба, но и к разным видам (конечно одной группы, например мукоровых). Таким образом, здесь открывается возможность междувидовых скрещиваний. Правда, большею частью здесь дело не доходит до настоящего слияния, но иногда такая гибридизация удается до конца, например у Neurospora (Dodge, 1928), некоторых Ustilaginales (Kniep, 1926, Goldschmidt 1928, Kämmerling, 1929), Phycomyces blakesleanus X Ph. piloboloides (Burgeff, 1914, 1915, 1928).
Вместе с тем способность ( + ) и (—) мицелиев реагировать друг с другом при попытках междувидовых скрещиваний позволила установить общий стандарт этих обозначений для ряда видов. Таким образом, среди мукоровых обозначения (+) и (—) не являются случайными для каждого отдельного вида, но устанавливаются по сравнению с теми первоначальными расами, какие получили эти отметки в первых исследованиях Блексли (1904 и след.). У некоторых мукоровых (например Absidia) копулирующие органы не совсем одинаковы, а один больше и другой меньше. Они могут быть отождествлены — первые с женскими и вторые с мужскими половыми органами; также и производящие их мицелии могут быть определены как женские и мужские. Сравнение с ними других строго изогамных мукоровых позволяет установить, что здесь обозначение (+) физиологически соответствует женским, а (—) мужским формам (рис. 41, 3). Понятно, что такая стандартизация возможна только в тех группах, которые имеют половые органы. У совершенно лишенных их базидиальных грибов обозначения (+) и (—) не могут быть приурочены к определенному полу. Исключением оказывается здесь гомоталличный Zygorhynchus heterogamus. У него более крупный гаметангий даёт половую реакцию с (+) расами гетероталличных видов, т. е. несет, таким образом, половой заряд (—) (Satina, 1930).
Мицелии (+) и (—), т. е. соответственно женские и мужские, гетероталличных грибов иногда отличаются, кроме половой функции, еще и по ряду других свойств. Такие явления известны у мукоровых. Здесь иногда они отличаются по отношению к различным сахарам (Костычев и Елиасберг, 1919), скорости роста: (+) обыкновенно имеет более мощный рост и т. д. Отмечается, далее, большее отложение каротина и жира в (+) мицелиях некоторых видов (особенно в гаметофорах) и т. п. Такие явления часто трактуются как появление вторичных половых признаков. Однако связь указанных отличий именно с половой диференциацией далеко не всегда доказана. Более подробные и планомерно проведенные в этом направлении исследования Сатиной и Блексли показали, что у большинства гетероталличных мукоровых наблюдается более сильная восстановительная способность (восстановление солей селеновой и теллуровой кислоты, перманганата калия и др.) именно у (+) мицелиев, т. е. женских. По Burgeff, это явление зависит от большего скопления у женских рас дубильных веществ. Так как большая восстановительная способность свойственна женским экземплярам и среди некоторых других групп растений и животных, то в исследованиях Сатиной и Блексли можно, пожалуй, видеть указание на определенные биохимические отличия протоплазмы того и другого пола (+ и —) у исследованных ими грибов.