Perisporiales, как и Plectascales, имеют вполне замкнутые плодовые тела (клейстокарпии), но отличаются тем, что сумки в них, обыкновенно находящиеся в небольшом числе, расположены не без порядка, а соединены в пучок, поднимающийся от нижней части плодового тела. Сумки обычно овальны; парафизы между ними не развиваются. Большинство относящихся сюда форм являются поверхностными паразитами на листьях, реже сапрофитами.
Сюда относятся следующие два главные семейства:
- Семейство Erysiphaceae. Мицелий бесцветный. Исключительно паразиты.
- Семейство Реrisporiасеае. Мицелий темноокрашенный. Паразиты и сапрофиты.
1. Сем Erysiphaceae
Это очень естественное и характерное семейство представлено исключительно облигатно паразитными видами, поселяющимися на листьях и других молодых частях различных растений, где их поверхностно развитой мицелий образует беловатый пушок, покрывающийся затем как бы мучнистым налетом от развивающихся конидий (явление, известное под именем мучнистой росы, откуда название Erysiphaceae — мучнеросные грибы, рис. 170).

Erysiphaceae известно около сотни видов, из коих многие в свою очередь разделяются на ряд форм, или биотипов, имеющих иногда и мелкие морфологические отличия или отличающихся только физиологически, по способности заражать то или иное растение. Кроме того, известно немало таких, у которых наблюдались только конидиальные спороношения. Однако последние построены здесь столь характерно, что не оставляют сомнения в их принадлежности к Erysiphaceae. Большинство видов распространено в умеренных широтах, где они нередко имеют массовое развитие.
Мицелий
Мицелий у Erysiphaceae бесцветный, сложенный из одноядерных клеток. Он рыхло стелется по поверхности эпидермиса пораженного органа, но в некоторых местах более плотно прикладывается к эпидермальным клеткам, образуя здесь большей частью особые, иногда слегка лопастные расширения, называемые аппрессориями. От них развиваются гаустории, прободающие стенку клетки эпидермиса и вздувающиеся на конце, достигнув ее полости. В некоторых случаях, например у Erysiphe graminis, гаустория получает форму лопастного тела, занимающего почти всю полость клетки хозяина (рис. 171, 2). Гаустории являются единственным органом питания гриба и у большинства представителей проникают только в клетки эпидермиса. В некоторые случаях, например у Uncinula salicis и некоторых других, они заходят глубже, пронизывая насквозь эпидермальную клетку и вздуваясь только в полости субэпидермальной клетки мезофилла. Несколько иначе развит мицелий у Phyllactinia. Здесь поверхностный мицелий совсем не развивает гаусторий, но дает короткие, большей частью 4-клетные ветви, которые проникают внутрь листа через устьица, и уже их конечные клетки образуют гаустории (рис. 171, 3). Таким образом, у Phyllactinia в отличие от других Erysiphaceae известная часть мицелия (упомянутые 4-клетные ветви) оказывается эндофитной. Еще дальше пошло дело в том же направлении у Leveillula, где вначале весь мицелий развивается эндофитно и лишь позднее наружу через устьица выходят ветви его, развивающиеся в поверхностный мицелий.

1 — Erysiphe communis;
2 — Erysiphe graminis;
3 — Phyllactinia suffulta.
Органы бесполого спороношения представлены конидиеносцами в виде коротких приподнимающихся ветвей поверхностно стелящегося мицелия, отчленяющих на концах в базипетальном порядке четки овальных бесцветных конидий (рис. 170). У Leveillula такие же конидиеносцы высовываются наружу через устьица (рис. 176).
Эти очень характерно построенные конидиальные спороношения получили название Oidium или Oidiopsis (последнее в том случае, если, как у Leveillula или Phyllactinia, на конце конидиеносца сидит только одна конидия, а не целая цепочка их). Конидии здесь, как и все другие клетки Erysiphaceae, как в мицелии, так и в клейстокарпиях, содержат по одному клеточному ядру.
Сумчатые плодовые тела (клейстокарпии) образуются также у всех видов поверхностно (об условиях образования их см. стр. Условия возникновения нескольких спороношений). В зрелом состоянии они имеют вид темных шариков, еле различимых невооруженным глазом. Развитие их начинается с образования половых органов в виде коротких боковых выростов одной или двух соседних гиф поверхностного мицелия. Женский отросток состоит всего из одной одноядерной клетки — оогония, или аскогона, а мужской, более тонкий, делится на две клетки: нижнюю — ножку и верхнюю — собственно антеридий; обе также одноядерные. Антеридий прикладывается к верхней части аскогона, оболочки на месте соприкосновения растворяются, и мужское ядро переходит в аскогон (рис. 172).

1 — 5 — оплодотворение и ранние стадии развития плодового тела;
5 — видна двуядерная клетка, развивающаяся далее в единственную сумку.
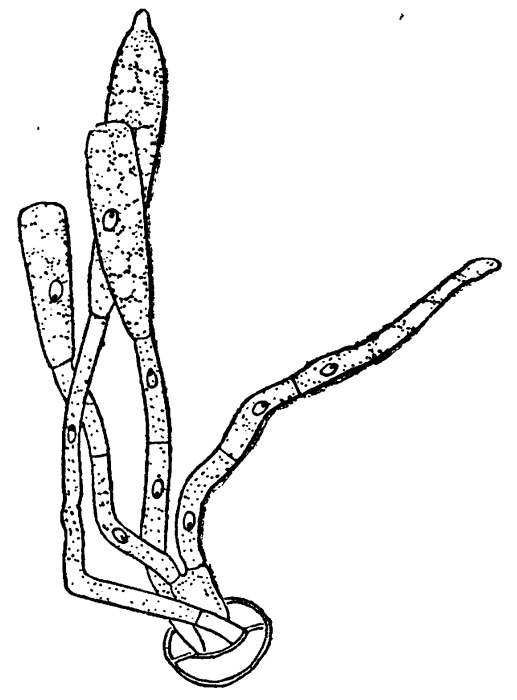
Согласно исследованиям прежних авторов (Герпер и др.), сейчас же за этим следует и слияние ядер, однако, согласно современным представлениям о половом процессе у высших грибов (см. стр. Половое воспроизведение, смена генераций и ядерных фаз у аскомицетов), такой немедленной кариогамии здесь происходить не должно, и ядра только сближаются друг с другом. Это представление находит подтверждение и в некоторых более новых, правда немногочисленных, исследованиях (Бессонов, 1914). После оплодотворения аскогон разрастается в многоядерный мешок, который затем делится поперечными перегородками на ряд клеток, из них одна, предпоследняя, оказывается в большинстве случаев двуядерной, а другие — одноядерными. У Sphaerotheca, где в плодовом теле имеется всего одна сумка, эта последняя образуется прямо из вышеуказанной двуядерной клетки, причем в ней происходит кариогамия и т. д. (рис. 172, 5). У таких родов, как Erysiphe или Phyllactinia, где сумок в плодовом теле несколько, из двуядерной клетки (или двуядерных, если их было несколько) образуются короткие аскогенные гифы, и уже на их концах формируются сумки (образования крючков здесь, повидимому, не происходит, но имеет место нормальная кариогамия и дальнейшее редукционное деление копуляционного ядра) (рис. 173).
а — более молодая стадия, видны аскогенные гифы;
b — более взрослая стадия, видны сумки, каждая с одним копуляционным ядром.
Несмотря на многочисленные исследования, посвященные половому процессу Еrуsiphaceae, многое здесь остается еще невыясненным. Так, прежде всего не ясно, каким образом у оплодотворенного аскогона с его дикарионом получаются снова одноядерные клетки и между ними одна или несколько двуядерных. С другой стороны, некоторые авторы, как Данжар, отрицают здесь оплодотворение в смысле слияния карпогона с антеридием (см. стр. Половое воспроизведение, смена генераций и ядерных фаз у аскомицетов).
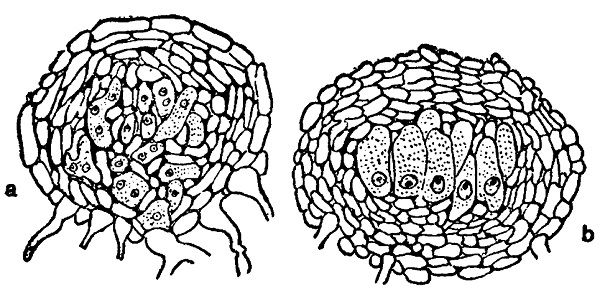
Одновременно с описанным развитием оплодотворенного аскогона происходит его обрастание гифами, берущими начало преимущественно прямо под основанием половых органов. Сначала они располагаются в один слой, а затем получают многослойное расположение и приобретают псевдопаренхиматический характер. Наружный слой их, сложенный из плоских табличатых клеток, буреет, его оболочки утолщаются, и он образует собственно оболочку клейстокарпия. Что же касается внутренних слоев, то они сохраняют нежные бесцветные стенки и образуют так называемое ядро плодового тела. Среди его псевдопаренхиматических элементов располагается пучок сумок (или одна сумка).
Некоторые клетки оболочки клейстокарпия дают выросты, развивающиеся в так называемые придатки (appendices), играющие, повидимому, известную роль в распространении плодовых тел, а вместе с тем по постоянству своего строения представляющие весьма важные систематические признаки. В некоторых случаях они мало чем отличаются от обыкновенных гиф мицелия и, стелясь по субстрату, переплетаются с ними (например у Erysiphe, рис. 174, а).

а — Erysiphe;
b — Microsphaera;
с — Uncinula;
d — прорастающий клейстокарпии Sphaerotheca, одна сумка со спорами.
У большинства других они имеют более специальное строение и идут свободно или в горизонтальной плоскости, или приподнимаясь кверху. Так, например, у Microsphaera придатки повторно вильчато разветвляются на концах на манер оленьих рогов (рис. 174, b), у Uncihula они загнуты на конце крючком (рис. 174, с). У Phyllactinia имеются придатки двух сортов: одни отходят от экваториальной линии клейстокарпия и расходятся горизонтально в виде прямых неразветвленных шипов с луковицеобразным расширением у основания; другие придатки на том же клейстокарпии отходят кверху от его полярной части и разветвлены в виде кисточек. Они выделяют слизистое липкое вещество, которое сливается в общую крупную каплю на полюсе плодового тела (рис. 175).

l — клейстокарпий в разрезе с придатками двух родов;
2 — цельный клейстокарпий, приподнятый на придатках, на верхнем полюсе — капля слизи.
При созревании клейстокарпии у форм с свободными придатками отрываются от субстрата. Этому способствует, с одной стороны, строение самой оболочки клейстокарпия, часто более тонкой с нижней стороны, вследствие чего при высыхании она впячивается внутрь и обрывает прикрепленные здесь гифы мицелия. С другой стороны, в этом отделении от субстрата имеют известное значение и придатки. Они гигроскопически изгибаются и, зацепляясь и склеиваясь друг с другом концами, соединяют плодовые тела в рыхлые комки, которые легко подхватываются ветром.
У Phyllactinia наблюдаются особенно любопытные приспособления в указанном направлении. Основные вздутия горизонтальных шипообразных придатков имеют здесь более тонкие стенки с нижней стороны и более толстые с верхней. Вследствие этого при высыхании нижняя сторона более сокращается (дает складки и т. д.), и в результате придаток поворачивается книзу. Так как это происходит более или менее одновременно во всех придатках, то в результате клейстокарпий приподнимается на них, как на ходулях (рис. 175, 2). В таком виде он легко сдувается ветром и, попав на другой субстрат, приклеивается к нему каплей слизи, выделяемой верхними кистевидными придатками. Благодаря указанному строению клейстокарпий, у большинства Erysiphaсеае отделяющийся от субстрата, является сам по себе органом распространения гриба. У Erysiphe и др., где придатки переплетаются с мицелием, отделение плодовых тел от субстрата происходит труднее и имеет место только после разрушения (сгнивания) мицелия. По указаниям Gossipovitch et Mladen (1929), приатки плодовых тел у Uncinula и др. служат не для рассеивания их, а для удержания на месте. Их кончики ослизнены и в сухую погоду приклеиваются к субстрату, но отклеиваются во время дождя.
Клейстокарпий прорастают обыкновенно только на другой год, и даже аскоспоры у некоторых видов образуются в сумках только после зимовки. Самое прорастание происходит таким образом, что сумки и внутренняя псевдопаренхима плодового тела взбухают, впитывая воду. Под их давлением оболочка плодового тела разрывается, сумки высовываются наружу и выбрасывают свои споры (рис. 174, d). Аскоспоры подхватываются ветром и, попадая на соответствующее растение, дают первое весеннее заражение его. Образующиеся здесь вскоре конидии служат для быстрого размножения гриба в течение лета. Они способны к немедленному прорастанию и быстро теряют свою всхожесть. У некоторых видов (Sphaerotheca pannosa, Uncinula spiralis) способна к зимовке и самая грибница в трещинах коры или в почках.
Условия развития Erysiphaceaе
По образу жизни все Erysiphaceae облигатные паразиты. Хотя их аскоспоры и конидии легко прорастают просто в капле воды, давая с узких концов своих ростковые трубочки, но получить дальнейшее развитие их в сапрофитных условиях до сих пор не удалось. В естественной обстановке в паразитных условиях развитию Erysiphaceae благоприятствуют сухость атмосферы и яркое освещение. Согласно исследованиям Hammarlund (1925) в сухом воздухе образование конидий происходит обильнее и всхожесть их больше. Такое же повышение всхожести наблюдается у конидии, образовавшихся на ярком свету, по сравнению с теми, которые развились в темноте или при пониженном освещении. Непосредственные наблюдения в природе также показывают, что особо сильное развитие Erysiphaceae имеет место в сухое лето или вообще в странах с сухим климатом (например в средней или передней Азии). Лишь для первых стадий инфекции они нуждаются, повидимому, в капельножидкой воде в виде росы или дождя. С другой стороны, отмечается очень сильное развитие некоторых Erysiphaceae в оранжереях и вегетационных домиках, где они нередко вызывают большие опустошения (например Sphaerotheca pannosa на розах и особенно Erysiphe graminis на злаках в вегетационных опытах).
Воздействие на ткани
По характеру воздействия на пораженные ткани Erysiphaceae, как и другие облигатные паразиты, характеризуются тем, что по крайней, мере на первых стадиях развития отношения между хозяином и паразитом напоминают скорее симбиоз, чем обычный паразитизм: клетки хозяина, даже те, в которые внедрилась гаустория гриба, не отмирают, а показывают иногда даже как бы известные признаки стимуляции в своей жизнедеятельности в виде некоторого разрастания, накопления питательных веществ и т. д. Листья клена, пораженные Uncinula aceris, значительно дольше сохраняют зеленую окраску осенью в пораженных, местах, чем в свободных от паразита.
Классификация Erysiphaceae устанавливается на основании строения клейстокарпиев. Главнешие роды следующие:
I. Клейстокарпии с одной сумкой.
1) Sphaerotheca. Придатки нитевидные, переплетающиеся с грибницей.
2) Podosphaera. Придатки не гибкие, свободно растущие вверх, вильчато разветвленные на концах.
II. Клейстокарпии с несколькими сумками.
3) Erysiphe. Придатки нитевидные, переплетающиеся с грибницей.
4) Leveillula. Клейстокарпии и их придатки, как у Erysiphe, но грибница первоначально эндофитная. Позднее из нее развивается и эктофитная грибница, а на ней образуются клейстокарпии.
5) Microsphaera. Придатки свободные, на концах вильчато разветвленные.
6) Uncinula. Придатки свободные, на концах закрученные спиралью или крючком.
7) Phyllactinia. Придатки двух родов: одни — отходящие экваториально, шиловидные, с вздутием у основания; другие — расположенные на вершине клейстокарпия, разветвленные кистевидно.
Наиболее важные и распространенные виды
Из представителей, имеющих особо важное значение или особо распространенных, заслуживают упоминания:
Sphaerotheca pannosa с двумя формами: f. persicae (на персиках) и f. rоsae (на розах). Очень важный вредитель названных растений.
Sph. mors uvae на видах Ribes. Этот паразит занесен в 1900 г. из Америки и теперь всюду распространен в Европе, являясь очень важным вредителем крыжовника.
Sph. castagnei (Sph. humuli или Sph. macularis) с очень большим количеством физиологических форм на разных растениях.
Podosphaera oxyacanthae с рядом форм на различных розоцветных (яблоня, айва, персик и др.).
Erysiphe gramirris с очень большим количеством форм, приуроченных к разным злакам.
Erysiphe labiatarum с большим количеством форм на различных губоцветных.
Erysiphe cichoriacearum — сборный вид с большим количеством форм на различных растениях, особенно на сложноцветных.
Erysiphe communis (Erys. polygoni — сборный вид, с массой форм, установлено 130) на растениях различных семейств.
Microsphaera alphitoides на листьях молодых побегов дуба; очень распространенный вид, до недавнего времени известный лишь в конидиальной стадии под названием Oidium dubium или Oidium quercinum; последние годы у нас наблюдается всюду обильное образование клейстокарпиев.
Uncinula necator (U. spiralis = Oidium Tuckeri) — серьезный вредитель винограда, вызывающий болезнь, известную под названием пепелицы. Гриб занесен из Северной Америки в 70-х годах прошлого столетия и первоначально был известен только в конидиальном состоянии (Oidium Tuckeri); сейчас обнаружены и клейстокарпии (в Крыму).
Uncinula salicis и U aceris — очень распространенные виды на ивах и кленах.
Leveillula taurica — повидимому, сборный вид на растениях из различных семейств. Огромного развития достигает в Малой и Средней Азии, где придает характерный вид пейзажу, покрывая листья растений мучнистым сероватым налетом.
Phylactinia suffulta — сборный вид на различных древесных и травянистых растениях.
Борьба с мучнеросными грибами, производящими иногда значительные опустошения в тех или иных культурах, облегчается их поверхностным положением на растении. Здесь наиболее применяют опыливание растений мелко раздробленной серой (серный цвет). На некоторые другие виды (например Sphaeroth. mors uvae) сильно действуют мышьяковистые соли.
Erysiphaceae представляют очень хорошо ограниченную группу специализированных паразитов, приспособившихся к эктофитному существованию. Филогенетически они связываются, как и остальные Perisporiales, с Plectascales. Что касается эволюции в пределах самого семейства Erysiphaceae, то некоторые, как Гейман, ставят здесь вначале Leveillula с ее эндофитным мицелием. Однако кажется более вероятным, что эндофитизм ее — вторичного происхождения и что в начале ряда нужно поставить такие формы, как Егуsiphe с их мало диференцированными придатками на плодовых телах. Отсюда эволюция могла идти в следующих основных направлениях:
вторичное приспособление к эндофитизму (Leveillula, отчасти Phyllactinia);
диференцировка придатков в свободные образования (Microsphaera, Uncinula, Phyllactinia).
Односумковые роды, Sphaerotheca и Podosphaera, являются параллельными: первая Erysiphe, а вторая — Microsphaera и, вероятно, приобрели этот признак вторично.
2. Сем Perisporiaceae
В отличие от Erysiphaceae, сем. Perisporiaceae ограничено весьма неясно и связывается постепенными переходами, с одной стороны, с Plectascales, а с другой — Sphaeriales и другими пиреномицетами. Сюда относятся преимущественно тропические и субтропические формы, в огромном большинстве эктофитные паразиты. Мицелий поверхностный, темноокрашенный, прикрепляющийся к эпидермису особыми отростками, так называемыми гифоподиями. Кроме того, у большинства развивается и внутренний мицелий, в виде ответвлений наружного проникающий в лист через устьица.
Этот внутренний мицелий бессцветный (рис. 177). Такой характер строения и роста мицелия встречается кое-где и в других группах аскомицетов (например Sphaeriales), особенно растущих в сырых тропических местообитаниях. В этих случаях, так же как и во многих других, поверхностный темноокрашенный мицелий разрастается радиально и более или менее равномерно во все стороны из общего центра и носит название астериноидного (греческое aster — звезда).

1 — разрез через лист с поверхностным темным мицелием и внутренним светлым;
2 — разрез листа с плодовым телом гриба.
На наружном мицелии образуются сумчатые плодовые тела строения, сходного с Erysiphaceae (рис. 177, 2). В некоторых случаях они содержат по одной только сумке (Balladyna), в других по нескольку, расположенных пучками. Из относимых сюда представителей можно указать еще Parodiopsis и Meliola. У последней плодовые тела очень похожи на Erysiphaceae, иногда с такими же придатками (М. furcata).
Сюда же нередко присоединяют грибы так называемой «черни», очень обычные у нас на листьях разных растений (тополь, слива, яблоня, малина и др.) и объединяемые часто в род Apiosporium (Capnodium или Fumago). По существу — это сапрофиты, питающиеся на листе выделениями тлей. Мицелий их поверхностный, темноокрашенный, без гаусторий. На нем описаны весьма различные спороношения. Во-первых, темные хдамидоспоры, на которые распадаются гифы; в сахаристых средах они размножаются почкованием (Dеmatium pullulans старых авторов). Во-вторых, указаны различные конидиальные спороношения: отдельные конидиеносцы, коремии, расширенные наверху чашевидно, пикниды двух сортов. Наконец, указываются и сумчатые спороношения, развивающиеся на особых темноокрашенных ветвящихся стромах. Согласно, некоторым современным данным здесь объединено несколько различных грибов разного систематического положения, но недостаточно, еще изученных. Самое название Apiosporium предлагается поэтому упразднить.




























