Согласно этой гипотезе половая тетраполярность объясняется таким образом, что гаплонты отличаются друг от друга по существу только одним половым фактором, т. е. в основе являются обычными биполярными (мужскими или женскими), но, кроме того, они содержат еще особый фактор самостерильности, который препятствует их соединению. Последнее наступает лишь в том случае, если эти факторы самостерильности различны. Таким образом, из каждой пары половых факторов Книпа {АВ, ab, Аb,аВ) один толкуется как собственно половой, а другой — как фактор самостерильности. В пользу такого рода представлений можно привести не только то, что они сводят дело к более привычным нам двум полам, но и то, что наличие факторов самостерильности довольно хорошо доказано у некоторых высших растений (например у отдельных рас табака) Корренсом. Повидимому, и у грибов они также обнаруживаются. Например, у Ascobolus magnificus, биполярно гетероталличного аскомицета, каждый односпоровый мицелий развивает и женские и мужские органы, но они не копулируют друг с другом, и поэтому апотеции в этом случае не развиваются. Если же встречаются мицелий (+) с мицелием (—), то копуляция, повидимому, происходит между мужскими и женскими органами разных мицелиев и дальнейшее развитие идет нормально (Gwynne-Vaughan and Williamson, 1932). Можно думать, что в основе указанных явлений лежат именно факторы самостерильности, одинаковые в органах односпорового мицелия и различные в разных мицелиях, обозначаемых как (+) и (—). Аналогичные данные имеются и относительно некоторых других аскомицетов.
Рубрика: Проф. Л. И. Курсанов «Микология»
Гипотеза Гартмана
Для объяснения такого сорта явлений возможно воспользоваться дополнительной к Книпу гипотезой, предложенной Гартманом (1929) и еще ранее для высших растений высказанной Корренсом (1907, 1913). Согласно ей как мужские, так и женские гаметы (соответственно гаплоидные ядра раздельнополых форм) содержат в себе обе половые потенции, но, кроме того, в них присутствуют особые факторы-реализаторы, действие которых сводится, повидимому, к подавлению противоположного пола. Эти факторы-реализаторы по своему проявлению соответствуют тем половым факторам, которые принимает Книп, но отличаются тем, что им придается количественное значение, тогда как у Книпа они отличаются качественно. В случае биполярного разделения полов факторы-реализаторы присутствуют в диплоидном ядре в количестве одной пары, а в случае тетраполярного разделения — в количестве двух пар (каждая пара при редукционном делении расщепляется). Обозначая по предложению Корренса мужскую и женскую потенции как А и G, а факторы-реализаторы соответственного пола — как ɑ и ɤ, мы можем представить диплоидное ядро биполярных в половом отношении форм следующим образом: [AG/ɑ AG/ɤ ]; при редукционном делении из него образуется по паре: [AG/ɑ] и [ AG/ɤ ]. Первая выявляется как мужская, так как присутствие реализатора ɑ подавляет в ней женскую потенцию; вторая, наоборот, выявляется как женская. В случае тетраполярных в половом отношении форм их диплоидное ядро должно получить обозначение: [AG/ɑ ɤ AG/ ɑ’ ɤ’ ]. При редукции из него могут выявиться или четыре гаплонта, сходные попарно: [AG/ɑ ɤ] и [AG/ ɑ’ ɤ’ ] — случай Aleurodiscus, или все четыре различные: [AG/ɑ ɤ ], [ AG/ɑ ɤ’ ], [AG/ɑ’ ɤ], [ AG/ɑ’ ɤ’ ] — случай Coprinus lagopus.
Присутствующие здесь в каждой гамете факторы-реализаторы обоих полов имеют неодинаковую мощность, что и определяет, в конце концов, ту или иную половую функцию. Таким образом, половая диференцировка сводится, согласно этим представлениям, к количественным различиям — к разнице, так сказать, половых потенциалов. В эту дополнительную гипотезу Гартмана укладываются и те явления, которые представляют известные затруднения для гипотезы Книпа. Гомо-гетероталлизм сводится к изменению напряжения (выравниванию) реализаторов пола. Относительная сексуальность объясняется таким образом, что копуляция наступает, если между копулянтами существует некоторая разница половых потенциалов, не меньшая определенного minimum’a = n. Если представить, что А имеет потенциал = — n/2, а В = + n/2, то А и В копулируют; если далее С имеет потенциал не менее чем 1,5 n, то он будет копулировать как с А, так не В. Таким же образом разностью потенциалов объясняется и различная сила половой реакции в разных случаях («половая жадность»). Копуляция гаплонтов, происшедших из разных плодовых тел, также проще объясняется различной высотой потенциала, а не качественными отличиями соответственных аллеломорфов, так как в последнем случае приходится допускать очень большое количество этих качественно отличных аллеломорфов в разных местах. Наконец, преимуществом указанных количественных представлений является и то, что они позволяют свести тетраполярный тип разделения полов к двум привычным нам началам (мужскому и женскому) аналогично (но, конечно, не тождественно) положительному и отрицательному электрическому потенциалу. При такого рода представлениях относительно половой диференцировки нужно только иметь в виду, что количественные изменения реализаторов происходят не постепенно, а скачками, или, что одно и то же, результаты этих изменений сказываются лишь при переходе за известный предел.
Гипотеза Книпа
Указанные необычные и с первого взгляда очень сложные явления таловой диференцировки представляют весьма большой интерес в общебиологическом отношении. Для объяснения их приходится строить гипотезы. Наиболее разработана из них гипотеза Книпа (1922, 1928, 1929). Согласно ей при обычном биполярном разделении полов во время редукционного деления ядра происходит расщепление в одной паре половых факторов. Таким образом, если диплоидное ядро, точнее его содержащую половые признаки часть, мы обозначим как Aa, то после редукции из него получатся четыре ядра: А, А, а, а, которые попарно тождественны и соответствуют А — мужским (—) и а женским (+). При тетраполярном разделении полов происходит расщепление в двух парах факторов, причем они расщепляются самостоятельно и, стало быть, находятся в разных хромосомах. Диплоидное ядро при этом мы должны обозначить как AaBb. При редукционном делении из него образуются четыре ядра, в которых половые факторы могут распределяться следующим образом:
- АВ, АВ, ab, ab на одних базидиях и Ab, Ab, аВ, аВ на других или
- АВ, АЪ, аВ, ab на одной базидии.
Первый случай мы имеем, например, у Aleurodiscus polygonis, где на каждой базидии обычно образуются споры только двух сортов, а на другой — двух других сортов. Второй случай мы имеем у Coprinus lagopus или Collibia velutipes, где на одной базидии образуются все четыре сорта спор.
Цитологически два указанных типа можно объяснить таким образом, что у Aleurodiscus первое деление ядра в базидии оказывается редукционным по отношению к половым свойствам. В результате его получаются два не сходных генетически ядра, например АВ и ab в одних базидиях или Ab и аВ в других. Делясь второй раз уже без редукции, они дают каждое по паре одинаковых
ядер. У Coprinus lagopus редукционным в отношении половых свойств оказывается второе деление. Поэтому получившиеся после первого деления два одинаковых ядра, при втором дают четыре неодинаковых (АВ, Ab, aB, ab). Впрочем, такая строгая связанность расхождения половых генов с тем или другим делением выдерживается не всегда. Позднейшими работами Книпа было обнаружено, что у Aleurodiscus некоторые тетрады базидиоспор (т. е. полученные каждая с одной базидии) оказываются диференцированными тетраполярно. Очевидно, в этом случае редукция по половым генам была передвинута на второе деление. Также и у Coprinus lagopus наблюдались отклонения от типичной для него тетраполярной диференцировки каждой тетрады в сторону биполярной. Это указывает на происшедшее в некоторых базидиях перемещение расхождения половых генов с второго деления на первое. Повидимому, такие перемещения могут вызываться в известной мере и внешними условиями развития базидии.
Согласно гипотезе Книпа полученные вышеуказанными путями гаплонты тетраполярных в половом отношении гименомицетов могут нормально соединяться с другими лишь в том случае, если они отличаются по обоим половым факторам, т. е. АВ копулирует с ab, или Ab копулирует с аВ, но в иных комбинациях копуляция не наступает. В результате копуляции в том и другом случае восстанавливается, в конце концов, снова диплоидное ядро Аа ВЬ.
Для объяснения половой реакции между спорами из разных плодовых тел Книп предполагает, что каждый половой фактор может существовать в нескольких видоизменениях (аллеломорфах). Если диплоидные ядра одного плодового тела мы обозначаем как AbВb, то на другом они могут соответствовать A1a1B1b1 и т. д. Из первого мы получили гаплонты АВ, Ab, aB, ab, и из второго — А1 В1, A1 b1, a1 B1, а1 Ь1. Они отличаются в обоих факторах и могут поэтому копулировать в любых комбинациях. В случае если видоизмененной является только одна пара факторов, то копуляция должна наступить лишь в половине возможных комбинаций. Например АаВb дает гаплонты АВ, Ab, aB, ab, a А1 а1 ВЬ — дает гаплонты А1В, А1b, а1В, а1b; каждый из этой четверки отличается обоими факторами только от двух гаплонтов другой четверки. Гипотеза Книпа хорошо согласуется с результатами многочисленных комбинаций гаплоидных мицелиев, которые были проведены как им, так и другими исследователями.
Однако имеется ряд явлений, которые не совсем укладываются в эту гипотезу в ее чистом виде. Это будут, например, явления, так называемого, гетеро-гомоталлизма и, особенно, относительной сексуальности. Гетеро-гомоталлизмом называется превращение формы, бывшей в начале раздельнополой (гетероталличной) в обоеполую (гомоталличную). У некоторых форм это превращение наблюдается столь часто, что стираются границы между гомоталлизмом и гетероталлизмом. Такое явление наблюдалось как у форм с биполярной сексуальностью (например Goprinus radians, Vandendries, 1924), так и у форм тетраполярных (например Coprinus micaceus, Vandendries, 1927). Явление относительной сексуальности известно особенно у водорослей, но встречается также и среди грибов. Оно заключается в том, что если мы имеем две группы гамет (или, общее, — гаплонтов) А и В, которые копулируют друг с другом и могут быть обозначены одни как мужские, другие как женские (или соответственно — и +), то существует еще третья группа С, которая копулирует как с А, так и с В, т. е. ведет себя как бы женская в одном случае и как бы мужская — в другом.
К той же категории принадлежит нередко наблюдаемое явление, что при комбинации мицелиев одного вида, различаемых как (+) и (—), наступает в разных случаях весьма неодинаковая количественно половая реакция. В одних случаях она еле заметна или совсем даже отсутствует, в других выражена очень сильно со всеми переходами. Это установлено, например, у Penicillium luteum (Derx, 1925), Absidia (Blakeslee, 1927) и др.
Биполярное и тетраполярное разделение полов
Для раздельнополых организмов мы привыкли различать только два пола: мужской и женский, соответственно (—) и (+). Это же явление наблюдается и у многих грибов; таковы, например, все изученные в этом отношении мукоровые; то же имеет место у аскомицетов, ржавчинных, большинства головневых и некоторых гименомицетов. Однако такое обычное биполярное разделение полов не является у грибов единственным и даже, пожалуй, преобладающим. В ряде других случаев имеется так называемое тетраполярное разделение полов; при его наличии устанавливаются четыре группы неодинаковых в половом отношении спор (и вырастающих из них мицелиев), так, что каждая группа реагирует только с одной из трех остальных. Это явление установлено у некоторых головневых (Ustilago zeae, Hanna, 1929; Ust. longissima v. macrospora, Bauch, 1930) и особенно распространено и даже, повидимому, преобладает среди гименомицетов. Иногда бывает, что уже на одной базидии образуются все четыре сорта спор (например Coprinus lagopus, Newton, 1926), иногда же споры одной базидии большею частью попарно одинаковы, но на другой соседней базидии развиваются две другие пары, не сходные с первыми (например у Aleurodiscus polygonis, Kniep, 1922). Нередко дело еще осложняется тем, что споры, взятые с одного плодового тела, реагируют в половом отношении со всеми спорами другого плодового тела того же вида, но взятого из другого местообитания. Такое явление наблюдается как у биполярных в пределах одного плодового тела видов (Coprinus comatus и Copr. Friesii, Brunswik, 1924), так и у тетраполярных (Coprinus fimetarius, Brunswik, 1924; Schizophyllum commune, Kniep, 1922, 1930).
Цитология и наследственность
Цитологической основой генотипического определения пола является редукционное деление. У низших грибов оно имеет место при прорастании зиготы, а у сумчатых и базидиальных при образование аско- и базидиоспор. У фикомицетов прямые цитологические данные в указанном направлении очень скудны. Однако генетический анализ потомства прорастающей зиготы позволяет сделать некоторые весьма вероятные заключения и о тех цитологических процессах, какие здесь происходили при прорастании. У Phycomyces blakesleanus из прорастающей зиготы вскоре образуется зародышевый спорангий (Keimsporangium), споры которого большею частью бывают разных знаков, но не в одинаковом числе. Наряду с этим появляются иногда такие зародышевые спорангии, которые содержат споры только одного знака. Наконец, среди раздельнополых спор попадаются в небольшом числе и нейтральные. Далее, в опытах Burgeff (1928) при скрещивании двух разновидностей Phycomyces, отличающихся друг от друга тремя парами признаков, в потомстве оказалось не восемь различных форм, как следовало бы ожидать, а только четыре (гибрид Аа ВЬ Сс; возможные гаплонты 1) ABC, 2) АВс, 3) Abс, 4) АЬС, 5) аВС, 6) аbC, 7) аВс, 8) abс).
Это последнее обстоятельство заставляет сделать следующие весьма вероятные предположения: все потомство зиготы сводится к одному ядру, претерпевающему редукционное деление на четыре. Что касается других очень многочисленных ядер, содержащихся первоначально в зиготе, то они, вероятно, отмирают. Четыре ядра зиготы, возникшие в результате редукционного деления первичного копуляционного ядра, диференцированы на (+) и (—). В дальнейшем они еще многократно делятся эквационным способом (не меняя, конечно, своего полового знака) и дают в конце концов ядра спор, число коих в спорангии Phycomyces достигает 70—80 тысяч. Споры в зародышевом спорангии, как правило, закладываются одноядерными (в отличие от обычного спорангия на мицелии, где они с самого начала многоядерны). Иногда в зародышевом спорангии споры залагаются как многоядерные и, если в них попадают и (+) и (—) ядра, то в результате из них получается нейтральный гетерокариотический мицелий. Размножение первых четырех гаплоидных ядер происходит не с одинаковой скоростью: иногда преобладает размножение ядер (+), иногда ядер (—), или имеет место дальнейшее отмирание ядер одного какого-нибудь знака. В результате в первичном спорангии получается преобладание спор какого-нибудь одного знака. У Мuсоr mucedo это явление пошло еще дальше. Здесь всегда в зародышевом спорангии имеются только споры одного пола: или (+), или (—) (Blakeslee, 1906).
Что касается сумчатых и базидиальных грибов, то здесь имеются многочисленные цитологические исследования относительно редукционного деления копуляционного ядра в сумке и базидии. В первой в результате трех делений, в конце концов, образуется почти всегда восемь гаплоидных ядер, а во второй в результате двух делений — четыре. Каждое из них, как правило, дает ядро споры. Поэтому одна сумка или одна базидия у гетероталличных форм должна нести споры разных полов в равных количествах. Это и наблюдается в действительности.
Обычно считается, что разделение генов при редукции осуществляется во время первого (гетеротипного) деления. Однако поскольку дело касается половых свойств у грибов, это оказывается не всегда так.
Например Dodge и Wilcox (1927, 1928), изучив ядерные деления в развивающейся сумке Neurosporasitophila, установили, что они все происходят одновременно и ориентированы в общем продольно. После третьего деления получается: 8 ядер и около них 8 аскоспор, расположенных в один ряд друг за другом. Если теперь изолировать эти аскоспоры в том порядке, как они находятся в сумке, и определить их половые свойства, то можно решить, при каком делении произошло разделение полов. Если при первом, то сверху должны лежать подряд 4 споры одного полового знака, а снизу — другого. Если при третьем — то споры (+) и (—) должны чередоваться через одну, если при втором, то должны чередоваться пары спор одного знака с парами другого. Обыкновенно наблюдается последний случай, что и указывает на расхождение половых генов при втором делении (рис. 42). Согласно данным Lindegren (1932) у видов Neurospora разделение полов может происходить как при первом, так и при втором делении в сумке, причем у Neurospora crassa это значительно чаще происходит при первом делении.

В базидиях Ustilago происходит два ядерных деления, ориентированных продольно, и самая базидия делится затем на четыре клетки, лежащие друг над другом. Если первое деление ядра вело за собой разделение половых генов, то указанные клетки базидии или образуемые ими базидиоспоры должны чередоваться попарно, если же это происходило при втором делении,— то они должны чередоваться по половым свойствам, через одну. Фактически чаще бывает второе. В других случаях, как в неразделенных базидиях гименомицетов, применим иной метод, о котором будет сказано ниже. В результате же его применения оказывается, что у одних гименомицетов разделение полов происходит при первом делении ядра в базидии, у других — при втором.
Гетероталлизм
Явление раздельнополости у грибов впервые установлено Блексли в работах 1904 и 1906 гг.
Оказалось, что у большинства видов изучавшихся им мукоровых на мицелии, выращенном из одной споры, не образуется зигот. Они развиваются лишь при встрече двух мицелиев, притом не всяких, а только некоторых, хотя и одинаковых по внешности, но отличающихся друг от друга в половом отношении. На месте соприкосновения их на том и на другом залагаются органы оплодотворения, и в результате их слияния образуются зиготы (рис. 41, 1).

1 — Phycomyces образование половых органов на месте встречи (+) и (—) мицелиев;
2 — схема образований гетерокариотического мицелия в опыте Burgeff;
S — схема копуляции гомоталличной Absidia (в середине) с гетероталличным (+) и (—) Mucor (справа и слева).
Так как самое оплодотворение у мукоровых грибов обыкновенно имеет строго изогамный характер, то здесь не приходится говорить о мужском и женском мицелиях; поэтому Блексли обозначил их умадно знаками: одни (+), а другие (—), так же, как и производящие их споры, а самое явление такой раздельнополости назвал гетероталлизмом. В дальнейшем раздельнополость была установлена и у некоторых таких мукоровых, где, как у Absidia и др., имеются довольно ясные различия в величине и строении копулирующих органов (рис. 34, 2).
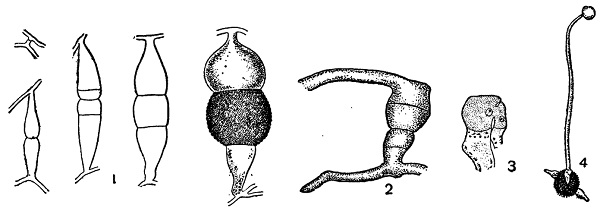
1 — Rhizopus nigricans;
2 — Absidia spinosa — гетерогамная копуляция;
3 — Endogone — гетерогамная копуляция одноядерных клеток;
4 — Мисоr — прорастание зигоспоры образованием зародышевого спорангия.
При этом при встрече соответственных двух мицелиев на одном из них всегда образуются большие (женские), а на другом меньшие (мужские) органы. В этом случае неопределенные обозначения мицелиев (+) и (—) могут быть заменены названиями женский и мужской, а обозначение гетероталлизм обычным — раздельнополость. Однако терминология Блексли столь укоренилась, что употребляется и сейчас почти постоянно для обозначения явлений распределения пола у низших растений (не только у грибов, но и у водорослей). Таким образом, гетероталлизм = раздельнополость, противоположное понятие гомоталлизм = обоеполость.
Гетероталлизм установлен среди всех основных групп грибов. Большинство оомицетов, повидимому, гомоталличны. Гетероталлизм среди них установлен только у немногих, например у нескольких видов Dictyuchus (D. sterilis) и некоторых Phytophthora, как Ph. Faberi и Ph. omnivora, причем у последней, наряду с гетероталличными, обнаружены также и гомоталличные особи. Наоборот, среди зигомицетов гетероталлизм, повидимому, преобладает. По крайней мере среди мукоровых, особенно подробно изученных в указанном отношении, около 60 видов оказалось гетероталличными и около 25 — гомоталличными.
Среди огромной группы сумчатых грибов гетероталлизм установлен до сих лор приблизительно у 25 видов, из коих половина приходится на представителей своеобразных Laboulbeniales. Из других групп сумчатых грибов гетероталличны: Taphrina epiphylla, Taphrina Klebhnü, Penicillium luteum, Ceratostomella coerulea, Neurospora sitophila, Ascobolus magnificus, Ascobolus carbonarius, Ascobolus stercorarius, Humaria granulata и некоторые другие. Нужно, однако, помнить, что аскомицеты только что начали изучать в этом отношении и, вероятно, в дальнейшем будет обнаружено немало еще гетероталличных форм среди них.
При культуре аскомицетов нередко приходится сталкиваться с таким явлением, что гриб в чистой культуре, несмотря на наилучшие как будто условия, упорно не дает сумчатых стадий, тогда как в естественной обстановке в природе они обычны у данной формы. Весьма вероятно, что в ряде случаев это различие имеет в своей основе гетероталлизм: в природе, где имеется разнородный споровой материал, легко может произойти встреча двух подходящих мицелиев, необходимых для развития сумчатых плодоношений, в чистой же культуре, выводимой, как правило, из одной споры, такие условия создаться не могут. Возможно, далее, что гетероталлизм лежит в основе некоторых Fungi imperfecti. В пределах этой огромной группы (около 25 тысяч видов) для многих видов установлено, что они являются только бесполыми стадиями тех или иных аскомицетов. Самая редкость сумчатых спороиошеиий в ряде случаев может быть объяснена их гетероталлизмом и тем, что разные полы имеют неодинаковое распространение. Наконец, возможно, что у некоторых форм один пол совершенно утрачен (или сделался крайне редким); тогда получается настоящий несовершенный гриб.
У базидиомицетов гетероталлизм особенно распространен. Среди головневых изучено в этом отношении больше пятнадцати видов, и все они оказались гетероталличными; весьма вероятно, что таковыми окажутся, если не все, то огромное большинство головневых.
У ржавчинных гетероталлизм установлен впервые только в 1927 г. (Craigie), но с тех пор изучено в этом отношении около 10 видов и 9 из них оказались гетероталличными. Также в самое последнее время обнаружен гетероталлизм у некоторых аврикуляриевых и дрожалковых (Auricularia Auricula Judae, некоторые Exidia).
Среди гименомицетов из изученных форм более пятидесяти оказались гетероталличными и только девять — гомоталличными. Вероятно, гетероталлизм здесь преобладает. Наконец, и у гастеромицетов, когда стали овладевать культурой и этих грибов, в последнее время также обнаружен гетероталлизм (у Sphaerobolus, Lorenz, 1933; Cyathus и Crucibulum, Fries, 1936).
Вообще можно сказать, что гетероталлизм для гаплоидной стадии, к которой, собственно, и относится это понятие, кажется более естественным и понятным, если считать, что определение половых свойств происходит генотипически при редукционном делении. В тех же случаях, где мы имеем явление гомоталлизма, или обоеполости, она может иметь своей основой или фенотипическое определение половой функции, не связанное с генами, локализирующимися в ядре (истинная обоеполость), или в основе ее может лежать так называемая гетерокариотичносць мицелия, т. е. содержание в нем одновременно как (+), так и (—) ядер. Первый тип, повидимому, более распространен. Он устанавливается, например, у большинства гомоталличных мукоровых: Sporodinia, Zygorhynchus, некоторых видов Mucor, Absidia. Это доказывается, между прочим, явлениями регенерации половых органов (до их слияния), что можно вызвать легким повреждением их. При этом каждый из них регенирует в мицелий, и на нем снова образуются копулирующие друг с другом половые органы. Таким образом доказывается, что даже ядра половых органов до их слияния не имеют окончательно фиксированной диференцировки в половом отношении. Что же касается второго типа гомоталлизма (гетерокариотичного), то таковой с достоверностью установлен у Phycomyces, где он получен экспериментально переливанием протоплазмы (с ядрами) из ( —) мицелия в (+) мицелий (Burgeff, 1915, рис. 41, 2). Такой мицелий обычно совсем не дает половых органов и не реагирует ни с (+), ни с (—), но иногда (очень редко) на нем одном образуются зиготы, вероятно, вследствие разделения или отмучивания разнородных ядер в один и другой из копулирующих органов. В естественной обстановке такие гетерокариотические мицелии должны возникать из первично многоядерной споры, содержащей и (+) и (—) ядра, или в результате анастомозов двух мицелиев. Судя по всему, такую же гетерокариотическую основу имеет гомоталличность Neurospora tetrasperma, где в отличие от близких N. sitophila и N. crassa (гетероталличных) аскоспоры залагаются как двуядерные образования (не сестринские ядра) (Dodge, 1927—1928; Wilcox, 1928) (рис. 40).

1 — аскоспора с дикарионом, ядра разных половых знаков, даны светлым и черным;
2 — гетерокариотический мицелий, клетки с одним или несколькими дикарионами; кое-где на конце дикарионы разъединяются и получаются одноядерные клетки.
Также у Pleurage anserina, гдe в сумке нормально залагается по 4 двуядерные споры, они вырастают в гомоталличный мицелий. Но иногда у этого вида вместо одной двуядерной споры развиваются две карликовые одноядерные. Эти последние уже прорастают одна в мицелий (+), другая в мицелий (—). Отсюда можно заключить, что гомоталлизм Pleurage anserina, берущий свое начало от двуядерных аскоспор, имеет в своей основе гетерокариотическое сложение мицелия (Dowding, 1931). Реакция, наблюдаемая при соединении двух разнородных мицелиев гетероталличных видов в случае гименомицетов, которые совершенно лишены половых органов, заключается в появлении анастомозов между их ветвями и переходе ядер из одной анастомозирующей клетки в другую. Таким образом создаются первые двуядерные клетки, которые, разрастаясь, и дают начало двуядерному мицелию. В других же случаях, где имеется настоящий половой процесс (например у мукоровых или Dictyuchus), реакция выражается в появлении на мицелиях половых органов, которые затем и сливаются друг с другом (рис. 41, 1). Любопытно, что такое образование половых органов происходит раньше, чем два мицелия придут в непосредственное соприкосновение. Таким образом, здесь имеется известное действие на расстоянии. У мукоровых это действие передается даже через пленку коллодия, отгораживающую два разнородных мицелия, и через воздух. Отсюда следует, что здесь имеет место возбуждающее действие веществ, с одной стороны, растворимых и диффундирующих через коллодий, а с другой — летучих (Burgeff, 1924).
Мицелии (+) и (—) реагируют друг с другом не только в том случае, когда они принадлежат к одному виду гриба, но и к разным видам (конечно одной группы, например мукоровых). Таким образом, здесь открывается возможность междувидовых скрещиваний. Правда, большею частью здесь дело не доходит до настоящего слияния, но иногда такая гибридизация удается до конца, например у Neurospora (Dodge, 1928), некоторых Ustilaginales (Kniep, 1926, Goldschmidt 1928, Kämmerling, 1929), Phycomyces blakesleanus X Ph. piloboloides (Burgeff, 1914, 1915, 1928).
Вместе с тем способность ( + ) и (—) мицелиев реагировать друг с другом при попытках междувидовых скрещиваний позволила установить общий стандарт этих обозначений для ряда видов. Таким образом, среди мукоровых обозначения (+) и (—) не являются случайными для каждого отдельного вида, но устанавливаются по сравнению с теми первоначальными расами, какие получили эти отметки в первых исследованиях Блексли (1904 и след.). У некоторых мукоровых (например Absidia) копулирующие органы не совсем одинаковы, а один больше и другой меньше. Они могут быть отождествлены — первые с женскими и вторые с мужскими половыми органами; также и производящие их мицелии могут быть определены как женские и мужские. Сравнение с ними других строго изогамных мукоровых позволяет установить, что здесь обозначение (+) физиологически соответствует женским, а (—) мужским формам (рис. 41, 3). Понятно, что такая стандартизация возможна только в тех группах, которые имеют половые органы. У совершенно лишенных их базидиальных грибов обозначения (+) и (—) не могут быть приурочены к определенному полу. Исключением оказывается здесь гомоталличный Zygorhynchus heterogamus. У него более крупный гаметангий даёт половую реакцию с (+) расами гетероталличных видов, т. е. несет, таким образом, половой заряд (—) (Satina, 1930).
Мицелии (+) и (—), т. е. соответственно женские и мужские, гетероталличных грибов иногда отличаются, кроме половой функции, еще и по ряду других свойств. Такие явления известны у мукоровых. Здесь иногда они отличаются по отношению к различным сахарам (Костычев и Елиасберг, 1919), скорости роста: (+) обыкновенно имеет более мощный рост и т. д. Отмечается, далее, большее отложение каротина и жира в (+) мицелиях некоторых видов (особенно в гаметофорах) и т. п. Такие явления часто трактуются как появление вторичных половых признаков. Однако связь указанных отличий именно с половой диференциацией далеко не всегда доказана. Более подробные и планомерно проведенные в этом направлении исследования Сатиной и Блексли показали, что у большинства гетероталличных мукоровых наблюдается более сильная восстановительная способность (восстановление солей селеновой и теллуровой кислоты, перманганата калия и др.) именно у (+) мицелиев, т. е. женских. По Burgeff, это явление зависит от большего скопления у женских рас дубильных веществ. Так как большая восстановительная способность свойственна женским экземплярам и среди некоторых других групп растений и животных, то в исследованиях Сатиной и Блексли можно, пожалуй, видеть указание на определенные биохимические отличия протоплазмы того и другого пола (+ и —) у исследованных ими грибов.
Результат полового воспроизведения у грибов
Конечным результатом полового воспроизведения у всех почти грибов является образование спор, служащих для размножения. У низших грибов это будут споры зародышевого спорангия, а у высших — аскоспоры и базидиоспоры. От чисто бесполых спор, как обычные спорангиоспоры и конидии, они существенным образом отличаются тем, что их образованию предшествует редукционное деление диплоидного ядра. При этом между вновь возникшими гаплоидными ядрами происходит распределение наследственных, зачатков (генов) и возникают новые комбинации их. Таким образом, получаются зародышевые клетки (споры) неоднородного в генетическом отношении характера, что и является по существу основным смыслом всякого полового процесса. Это можно констатировать по отношению к разным свойствам и всего лучше к распределению полов, так как здесь имеется наибольшее количество данных. У раздельнополых форм разделение полов происходит именно в сумках, базидиях, или зародышевых спорангиях, тогда как все споры так называемого обычного бесполого спорангия, или конидиеносца, оказываются в этом совершенно однородными (подробнее см. гл. VI).
Кроме этих чрезвычайно существенных генетических особенностей, споры, происшедшие половым путем, отличаются от обычных бесполых и способом своего развития. Споры зародышевого спорангия закладываются, насколько это известно, одноядерными, тогда как бесполые спорангиоспоры оказываются обычно с самого начала многоядерными. Далее, сумки и аскоспоры, которые раньше по учению Брефельда ставились в параллель с спорангиями низших грибов, отличаются от них всем ходом своего развития: во-первых, кариогамией и редукционным делением в сумке, во-вторых, тем, что аскоспоры в ней образуются путем свободного образования, тогда как спорангиоспоры — путем раскалывания всей протоплазмы спорангия на отдельные участки, в-третьих, наконец, тем, что аскоспора залагается одноядерной (за очень редкими исключениями, как Neurospora tetrasperma, рис. 40), хотя в дальнейшем вторично нередко делается
многоядерной и даже многоклеточной, тогда как спорангиоспоры, как правило, первично многоядерны. Также и базидия, которая, по учению Брефельда, сравнивалась с конидиеносцем, отличается от него такими же особенностями. Здесь также характерны кариогамия, редукционное деление и первичная одноядерпость базидиоспор — черты, которые совершенно отсутствуют у конидиеносца при всем иногда наблюдаемом внешнем сходстве его с базидией.
1 — аскоспора с дикарионом, ядра разных половых знаков, даны светлым и черным;
2 — гетерокариотический мицелий, клетки с одним или несколькими дикарионами; кое-где на конце дикарионы разъединяются и получаются одноядерные клетки.
Половое воспроизведение, смена генераций и ядерных фаз у базидиомицетов
Базидиомицеты зашли значительно дальше по пути утраты полового акта. Настоящие половые органы здесь совсем неизвестны, и оплодотворение заменяется апогамическим соединением клеточных ядер в дикарион или в результате соединения двух гаплоидных вегетативных клеток, или даже иногда в пределах одной, просто в результате деления ее ядра. В том и в другом случае первые клетки, содержащие дикарион, разрастаются затем в целый дикариофитный мицелий, гомологичный аскогенным гифам аскомицетов, но отличающийся тем, что он имеет самостоятельное существование и обычно преобладает в цикле развития базидиального гриба (рис. 5). Базидии закладываются или непосредственно на таком мицелии, или на особых плодовых телах, образованных им. (Базидиальное плодовое тело, сложенное нацело из диплоидного мицелия, отличается от сумчатого, которое, как мы видели выше, представляет образование двойственной морфологической природы, отчасти гаплоидной, отчасти диплоидной (см. Аскомицеты). )

1 —у фикомицетов;
2 — у аскомицетов;
3 — у базидиомицетов.
Тонкий контур — гаплоидная фаза, жирный—диплоидная фаза.
При развитии базидий происходит кариогамия дикариона, и получившееся диплоидное ядро четыре, давая гаплоидные ядра для четырех базидиоспбр, экзогенно образующихся на базидии (рис. 39, 2). Из базидиоспор вырастает гаплоидный, или первичный, мицелий, сложенный из одноядерных или иногда вторично многоядерных клеток, но без парной ассоциации ядер. На нем указанным выше образом опять образуется диплоидный, или вторичный, мицелий. В частности, у головнехвых грибов (Ustilaginales) первичная двуядерность осуществляется часто слиянием двух базидиоспор, иногда даже еще сидящих на базидии (Tilletia, рис. 39, 2). Из них затем вырастает дикариофитный мицелий, заканчивающий свое развитие образованием хламидоспор, из которых затем прямо вырастают базидии. Таким образом, здесь гаплоидная фаза до крайности сокращена, будучи представлена только базидиоспорами, вся же жизнь гриба проходит в диплоидном состоянии. У некоторых других головневых базидиоспоры могут несколько разрастаться, размножаться почкованием и даже давать небольшой гаплоидный мицелий, но это состояние большею частью непродолжительно и скоро заменяется диплоидным. У ржавчинных грибов (Uredinales) гаплоидная фаза представлена в большинстве случаев первичным одноядерным мицелием, на котором образуются сначала особые спороношения — спермогонии. Затем в залагающихся на нем следующих спороношеииях — эцидиях — появляются первые двуядерные клетки в результате попарного слияния двух соседних. С них начинается диплоидная фаза, все клетки которой, как правило, двуядерны. Она здесь обильно размножается бесполым путем. На ней образуются сначала эцидиоспоры; из них вырастает дикариофитный мицелий, развивающий уредоспоры; последние в свою очередь могут дать новый мицелий с уредоспорами и т. д. Наконец, на таком диплоидном мицелии развиваются телейтоспоры, в которых происходит слияние дикариона в диплоидное ядро. Из телейтоспор прямо вырастают базидии, причем диплоидное ядро делится редукционным способом и этим восстанавливается гаплоидная фаза.

1 — развитие базидии гименомицета, а — d — последовательные стадии;
2 — Tilletia — базидия с парной копуляцией базидиоспор;
3 — Hypochnus:
а — развитие базидий;
b — вторично двуядерная базидиоспора;
с — ее прорастание двуядерным мицелием.
У высших базидиальных грибов наблюдается в общем такая же смена фаз. Из базидиоспоры вырастает первичный гаплоидный мицелий, который может иногда размножаться бесполыми конидиями, но в большинстве случаев имеет непродолжительное существование. Из него вырастает диплоидный мицелий, берущий свое начало из первичных двуядерных клеток; получающихся большею частью в результате анастомозов между ветвями двух разных мицелиев (гетероталлизм, см. стр. 48 и след.), реже — одного. Обыкновенно он имеет более мощное развитие и часто оказывается даже многолетним. На образованных им плодовых телах развиваются базидии с базидиоспорами. В некоторых случаях гаплоидное состояние у таких базидиомицетов крайне укорочено. Например, у Hypochnus terrestris базидиоспора, еще сидя на базидии, делит свое ядро на два и при прорастании сразу дает двуядерный диплоидный мицелий (рис. 39, 3). Таким образом, фактически здесь гаплоидное состояние почти совершенно выпадает из цикла развития.
Половое воспроизведение, смена генераций и ядерных фаз у аскомицетов
Среди аскомицетов наиболее просто половое воспроизведение осуществляется у Protascales. Например, у Eremascus fertilis две рядом лежащие клетки мицелия дают около разделяющей их поперечной перегородки короткие одноядерные выросты, которые и сливаются друг с другом. Копуляционная клетка, или зигота, не превращается прямо в спору, как у низших грибов, а продолжает развиваться дальше, причем ее копуляционное диплоидное ядро делится три раза (происходит редукция числа хромозом), и около образовавшихся восьми ядер формируются свободно лежащие восемь спор (рис. 35, 1). Таким образом, здесь зигота прямо вырастает в сумку. Единственным диплоидным состоянием является копуляционная клетка, и ее диплоидное ядро сейчас же претерпевает редукцию при своем первом делении. Поэтому здесь, как и у низших грибов, по существу представлено только гаплоидное состояние. То же имеется у ряда других Protascales. У некоторых дрожжей, например у Schizosaccharomyces octosporus, сливаются при образовании сумки две свободно живущие клетки, т. е. наблюдается явление, которое можно назвать хологамией. Дальнейшее развитие происходит таким же образом, как у Eremascus (рис. 35, 1). Следует отметить, что здесь, как и у низших грибов, часто собственно оплодотворения, т. е. слияния клеток, не наступает, и сумка развивается партеногенетически из одной клетки; при этом, понятно, в ней не происходит слияния двух ядер и не наблюдается редукции ее ядра (гаплоидного) при образовании спор. Так обстоит, например, дело у некоторых видов Endomyces (например End. decipiens и др.) и также у многих дрожжей, в том числе у обыкновенных культурных пивных дрожжей (Saccharomyces cerevisiae).

1 — Eremascus fertilis:
a — d — последовательные стадии;
d — аск с 8 аскоспорами.
2 — Schizosaccharomyces octosporus — d — аск с 8 аскоспорами.
3 — Nadsonia fulvescens — гетерогамная копуляция с образованием сумки из выроста большей (женской) клетки.
У некоторых дрожжей и близких к ним грибов описано своеобразное извращение смены ядерных фаз в цикле развития. Например, у Saccharomycodes Ludwigii в сумке формируются четыре аскоспоры, которые при прорастании попарно копулируют и размножаются дальше почкованием уже в диплоидном состоянии. При этих условиях понятно, что сумка здесь образуется без копуляции, прямо из вегетативной диплоидной клетки. По новейшим данным такое извращение смены ядерных фаз (вся жизнь — в диплоидном состоянии и только аскоспоры гаплоидны) встречается и у многих других дрожжевых грибов. При этом в ряде случаев отмечается, что преобладание диплоидной или гаплоидной фазы может вызываться условиями культуры (Manuel, Renaud, 1937).
Все остальные сумчатые грибы, которые можно назвать Euascomycetes, отличаются от Protascales тем, что зигота у них не превращается прямо в сумку, а дает выросты, называемые аскогенными гифами, и уже на их концах формируются сумки. В связи с этим слияние ядер копулирующих клеток запаздывает и происходит только в развивающейся сумке. Подобное запоздание кариогамии не является редкостью и среди низших грибов, где иногда половые ядра не сливаются в зиготе (покоящейся споре) до самого момента ее прорастания. Аналогичное явление наблюдается и у некоторых Protascales, например у Nadsonia. У этого дрожжевого грибка имеется анизогамная копуляция, и содержимое меньшей клетки переходит в большую (рис. 35, 3). Однако эта последняя не превращается прямо в сумку, а дает вырост, куда переходят оба ядра. Только здесь они сливаются, а вырост затем развивается в сумку. Однако все эти явления у Euascomycetes пошли значительно дальше: здесь половые ядра не только не сливаются тотчас же после слияния клеток, но, сближенные попарно в дикарион, размножаются.
Более ясное представление даст здесь рассмотрение реального примера, каким можно взять Pyronema confluens, образцово изученную в этом отношении Claussen’om (1912). Женский половой орган, называемый здесь архикарпом, состоит из двух клеток: нижней, вздутой, — аскогона — и верхней, цилиндрической, — трихогины. Обе они, как и вообще клетки у Pyronema, многоядерны. Рядом с архикарпом развивается мужской орган — антеридий в виде довольно массивной цилиндрической клетки, также многоядерной. Такие половые органы залагаются обыкновенно сближенными группами из нескольких пар, образуя так называемую розетку рис. 36, 2). В известный момент кончик антеридия плотно срастается с концом трихогины, разделяющая их перегородка растворяется, и антеридиалыюе содержимое вместе с несколькими десятками своих ядер переливается в трихогину собственное содержимое которой к этому времени дегенерирует. Затем в поперечной перегородке в основании трихогины появляется на короткое время отверстие, и через него антеридиальные ядра переходят в аскогон. Там они сближаются попарно с соответствующими женскими ядрами, но вопреки более ранним указаниям Harper’a (1900) и др. не сливаются с ними (рис. 36, 2). Затем из аскогона вырастает от 10 до 20 мешковидных выростов, которые начинают вытягиваться и ветвиться, образуя аскогенные гифы. В них пеоеходят попарно сближенные ядра (дикарионы) и размножаются здесь, делясь каждая пара одновременно и с параллельным расположением осей деления (конъюгированное деление), так что получаются новые пары, опять составленные из потомка мужского и женского ядер (рис. 36, 3). Аскогенные гифы делятся затем поперечными перегородками на клетки, из которых нижние (ближайшие к аскогону) содержат по несколько пар ядер, а верхние — по одной паре. Наконец, на концах аскогенных гиф начинают развиваться сумки. При этом конечная клетка характерным образом загибается в крючок, и оба ядра дикариона одновременно делятся, так что получаются две пары их. Одна из них лежит на месте перегиба крючка, а из другой одно ядро попадает в кончик, а другое в основание его. Вскоре появляются две перегородки, отграничивающие среднюю двуядерную клетку, которая является материнской клеткой сумки (рис. 36, 4 и 37, 1).

1 — группа половых органов в начале развития плодового тела;
2 — момент оплодотворения; содержимое антеридия ♂ перешло через трихогину в аскогон ♀, где замечаются пары ♂ и ♀ ядер;
3 — развитие аскогенных гиф;
4 — схема образования крючков и асков на концах аскогенных гиф. Ядра заштрихованные и бесцветные — соответственно мужские и женские.
Два ядра ее дикариона (соответственно мужское и женское) сливаются, и получившееся диплоидное копуляционное ядро делится редукционным способом, восстанавливая гаплоидное число хромозом (у данного вида их 12). За первым делением следует второе и третье, и около восьми образовавшихся ядер в разросшейся к тому времени сумке формируется восемь аскоспор (рис. 37). Их развитие происходит чрезвычайно характерным для всех аскомицетов путем свободного образования, в подробностях изученного Harper’om (1897 и след.) и др. После третьего митоза центрозомы, расположенные на противоположных сторонах вновь образовавшихся ядер, отодвигаются от них, причем ядра оттягиваются в носик. Затем лучистые фигуры, расходящиеся от каждой центрозомы, начинают загибаться в сторону ядра, принимая вид как бы колокола. Нарастая своим свободным краем, он смыкается, в конце концов, на противоположном полюсе. Таким образом, из общей протоплазмы сумки вырезается восемь участков, около восьми ядер. На границе их выделяется твердая оболочка, и они превращаются в аскоспоры (рис. 37).

1 — h — крючок и двуядерный молодой аск, а — аск с одним копуляционным ядром;
2, 3 и 4 — первое, второе и третье деление ядер;
5 — формирование аскоспор.
Что касается оставшихся одноядерными конечной и основной клеток крючка, то они затем обыкновенно сливаются друг с другом, а получившаяся таким путем новая двуядерная клетка или прямо вырастает в новый крючок, или дает более длинную ветвь, также на конце крючковидно загибающуюся. В результате этого процесса увеличивается число сумок на аскогенных гифах.
Наиболее характерная особенность описанного у Pyronema процесса развития заключается в том, что два существенных момента оплодотворения— слияние протоплазмы клеток (плазмогамия) и слияние ядер (кариогамия), весьма сближенные у других организмов, в том числе и у большинства низших грибов, здесь весьма раздвинуты, и между ними вставлено длительное развитие аскогенных гиф, в течение которого ядра размножаются в состоянии дикариона, соответствующего каждый по существу одному диплоидному ядру (общее число — хромозом в дакарионе, сложенном из двух гаплоидных ядер, конечно, диплоидное). Таким образом, этот участок цикла развития от плазмогамии до кариогамии гомологичен диплоидной фазе и по характерной особенности своего ядерного аппарата называется часто дикариофитом (также синкариофитом). Ему противополагается гаплоидная фаза, начинающаяся с аскоспор и через весь вегетативный мицелий доходящая до начального этапа оплодотворения, или плазмогамии.
Указанные черты являются общими для всех Euascomycetes. Особенность Pyronema, свойственная, впрочем, и большинству других аскомицетов, заключается в том, что здесь благодаря многоядерности половых органов происходит образование сразу многих дикарионов, т. е. имеется множественное оплодотворение, аналогичное, например, тому, что мы видели у Mucoraceae. Затем как для Pyronema, так и для других Euascomycetes характерно, что одновременно с описанным процессом полового воспроизведения около или вокруг половых органов и развивающихся аскогенных гиф разрастаются ветви вегетативного мицелия, которые, переплетаясь в плектенхиму, слагаются в тело, имеющее определенную для каждой группы форму и называемое сумчатым плодовым телом, или аскокарпом. В соответствии с указанным развитием оно представляется образованием двойственной морфологической природы. Главная масса его состоит из плектенхимы, образованной из мицелия, т. е. принадлежит к гаплоидной генерации, а внутри его проходят аскогенные гифы, представляющие, как было выяснено, диплоидную генерацию.

1 и 2 — Ascobolus carbonarius;
2 — начало развития аскогенных гиф из двух клеток аскогона;
3 — Ascobolus magnificus;
4 — PeniciIlium;
5 — Sphaerotheca;
6 — Polystigma;
7 — Rhizina undulata, развитие аскогенных гиф из ряда клеток архикарпа;
m — трихогина, к — конидия; ♀ — архикарп, ♂ — антеридий.
У многих аскомицетов строение половых органов и половое воспроизведение сходны во всех существенных чертах с Pyronema (Ascodesmis nigricans, Cubonia brachyasca, Ascobolus vinosus), у других же имеются известные отличия, причем даже близкие формы оказываются иногда далеко не сходными друг с другом в этом отношении. Например, у Ascobolus magnificus женский орган, или архикарп, представляет многоклетную ветвь. Ее конечный участок из десяти и более сравнительно узких клеток представляет трихогину, а нижележащая широкая многоядерная клетка функционирует как аскогон, т. е. из нее после оплодотворения развиваются аскогенные гифы (рис. 38, 3). То же имеется у Ascobolus carbonarius, но только у него в качестве мужского органа функционирует особая конидия (рис. 38,1,2). Она не спадает с конидиеносца, и к ней прикладывается кончик многоклетной трихогины (такое развитие антеридия в виде конидии является здесь, вероятно, вторичным явлением). У простейших форм, как некоторые Plectascales (Gymnoascus, Penicillium) или некоторых Pyrenomycetes (Gnomonia, Chaetomium) мужские и женские органы только слегка отличаются по своей величине, представляя спирально закрученные короткие ветви, большею частью многоклетные в зрелом состоянии (рис. 38, 4). У некоторых других Pyrenomycetes, как, например, у Venturia, женский орган состоит из спирали аскогона и прямой многоклетной трихогины, которая прикладывается к конечной клетке прямой мужской ветви (антеридию).
В приведенных примерах клетки половых органов многоядерны. При оплодотворении (цитогамии или плазмогамии) антеридий переливает свое содержимое в конечную клетку трихогины, оттуда оно через частичное растворение поперечных перегородок достигает аскогона, и там многочисленные мужские ядра попарно ассоциируются с такими же женскими ядрами. Таким образом, и здесь, как и у Pyronema, происходит множественное оплодотворение (точнее, множественное образование дикарионов). В некоторых других случаях половые клетки одноядерны, как, например, у Magnusia nitida или у всех Erysiphaceae. Последние отличаются еще тем, что у них архикарп не имеет трихогины, так что аскогон непосредственно прикладывается к антеридию (рис. 38, 5). В случаях одноядерности половых клеток, конечно, образуется при оплодотворении только один дикарион, который в дальнейшем размножается.
У многих сумчатых грибов оплодотворения (цитогамии) не происходит; при этом или антеридий все-таки развивается (Lachnea stercorea), или чаще его совсем не образуется. Тем не менее аскогенные гифы берут начало из аскогона, в котором замечаются парно ассоциированные ядра. Они могут получаться различным путем: в результате перехода ядер из соседних клеток архикарпа, или вегетативных гиф, или соответственной перегруппировкой внутри одной многоядерной клетки аскогона. Первое наблюдается, например, у Polystigma rubrum, где в одноядерную клетку аскогона через отверстие в перегородке переходит одно ядро нижележащей много ядерной клетки (эта клетка называется также антеридиальной, но очевидно, что оплодотворяющая функция у нее вторичная) (рис. 38, 6). У Rhizina undulata, где аскогенные гифы развиваются из нескольких друг над другом лежащих клеток аскогона, в них переходят ядра как из верхних, так и из нижних клеток (рис. 38, 7). Такой же переход имеет место, например, у Ascobolus citrinus, Ascobolus
furfuraceus и, вероятно, у многих других. Парная ассоциация ядер в пределах одной многоядерной клетки аскогона имеется, например, у Нumaria granulata, Humaria anceps, Lasiobolus pulherrimus, Thecoteus Pelletierii — и, вероятно, у многих других. Наконец, известны случаи, когда и женский орган не развивается или во всяком случае не функционирует. Так, у Ophiobolus graminis хотя на молодых стадиях и закладывается архикарп с трихогиной, но в дальнейшем он отмирает. На его месте вырастают недиференцированные специально одноядерные клетки, которые попарно сливаются, давая двуядерные, и из них вырастают аскогенные гифы. У Humaria rutilans, многих Helvellaceae и некоторых других архикарп, повидимому, совсем не закладывается, а на известной стадии происходит переход клеточных ядер из одной вегетативной клетки в другую. Получающиеся в результате этого чисто апогамического процесса двуядерные клетки развивают далее аскогенные гифы. Термин «апогамия» употребляется здесь в широком первоначальном смысле для обозначения процесса, заменяющего настоящее утраченное оплодотворение, причем, однако, сохраняется смена морфологических генераций, или ядерных фаз. Такому пониманию соответствует почти целиком термин Винклера (1908) «псевдомиксис» или термин Гартмана (1909) «автомиксис», термин же «апогамия» употребляется ими в более узком смысле как часть понятия апомиксис, т. е. такого размножения, когда не происходит никаких клеточных или ядерных слияний. При этом апогамия трактуется как такой случай апомиксиса, когда из вегетативной клетки гаметофита вырастает спорофит, т. е. имеется смена морфологических генераций. Это более узкое толкование апогамии, нередко принимаемое в новейшей, особенно немецкой, литературе, может войти как часть в принятое здесь более широкое понимание.
Существенно отличаются от других аскомицетов в разбираемых здесь отношениях Exoascales и Laboulbeniales. У некоторых Exoascales (единственный род Taphrina) аскоспоры, выпав из сумок, попарно копулируют друг с другом и дают сразу диплоидный (синкариофитный) мицелий. На нем затем развиваются сумки, где происходит кариогамия. Таким образом, в противоположность другим сумчатым грибам здесь преобладает диплоидная фаза, а гаплоидная представлена только аскоспорами (нередко предварительно они размножаются, почкованием в гаплоидном состоянии). Аналогичное извращение соотношений ядерных фаз в цикле развития наблюдается также у некоторых дрожжевых грибов, например у Saccharomycodes Ludwigii, смотрите выше.
Главной особенностью Laboulbeniales является мужской половой аппарат, представленный здесь не антеридием, прикладывающимся к женскому органу, а самостоятельными клетками — спермациями, которые иногда развиваются даже на других экземплярах (раздельнополость) и переносятся на трихогину женского полового органа; здесь они, вероятно, переливают в нее свое содержимое, переходящее затем дальше вниз до аскогона. Возможно, что мужская половая функция спермациев, по общему строению и развитию сходных с конидиями, является здесь вторичной; подобно тому как это намечается у Ascobolus carbonarius.
Из приведенных примеров видно, что при всем разнообразии в подробностях полового воспроизведения у Euascomycetes в основном оно состоит в следующем. В наиболее типичных случаях имеется оплодотворение специального женского органа — архикарпа — мужским — антеридием. За этим первым этапом оплодотворения (цитогамией, или плазмогамией) не следует непосредственно слияние половых ядер (кариогамии), а из аскогона развиваются сначала аскогенные гифы, куда переходят сближенные половые ядра (дикарион); они размножаются еще в них и, наконец, сливаются в развивающейся сумке, что представляет заключительный момент полового процесса (кариогамия). В ряде случаев эта первоначальная картина видоизменяется главным образом в том направлении, что типичного оплодотворения женского органа мужским не происходит. При этом иногда мужской орган совсем не развивается, а иногда отсутствует и женский. Однако всегда в известный момент развития появляются дикарионы, происходит развитие аскогенных гиф и, наконец, кариогамия в развивающихся сумках.
Способ и место апогамного появления первых дикарионов у разных представителей не одинаковы. Иногда переходит в клетку аскогона ядро другой, рядом с ней лежащей и принадлежащей к той же ветви архикарпа, иногда просто в пределах многоядерного аскогона происходит перегруппировка его ядер в дикарионы. В том и другом случае центром образования аскогенных гиф является все-таки аскогон. Но иногда и он не функционирует в указанном направлении и даже совсем отсутствует. Тогда дикарионы возникают в результате соединения ядер двух вегетативных клеток, и из них вырастают аскогенные гифы. Все эти явления представляют различные способы замены исчезающего нормального оплодотворения, причем основной момент полового акта — смена ядерных фаз — обязательно сохраняется. Их можно назвать явлениями апогамии, которая, таким образом, оказывается широко распространенной у аскомицетов и, очевидно, возникает самостоятельно в отдельных филогенетических рядах их, чем и объясняется разнообразие наблюдаемых здесь апогамических процессов и несходство в этом отношении даже между близкими формами.
Изложенное толкование полового воспроизведения у аскомицетов сейчас наиболее принято. В основании его лежит исследование Claussen’a о Pyronema (1912). Однако существуют на этот предмет и другие взгляды. По представлению Наrреr’а, занимавшегося этими вопросами еще в конце XIX в., за слиянием половых клеток сейчас же следует и слияние их ядер, так что в аскогенных гифах содержатся уже диплоидные ядра. При их слиянии в сумке должно получиться тетраплоидное ядро (с четверным количеством хромозом). Возвращение к гаплоидным ядрам, содержащимся в аскоспорах, может произойти здесь в результате двойной редукции: при первом делении число хромозом уменьшается до диплоидного числа, а вероятно при третьем — до гаплоидного (это последнее деление получило даже специально название брахимейозис — укороченная редукция). Такие взгляды, кроме Наrреr’а до самого последнего времени защищаются особенно Gwynne-Vaughan (последняя работа 1937 г.). Однако они не могут считаться доказательными. Не говоря уже об общей довольно малой вероятности такого рода смены ядерных фаз (гаплоидная — диплоидная — тетраплоидная), совершенно неизвестной для других организмов, и сама цитологическая база, подводимая сюда в виде брахимейозиса, оказалась более чем сомнительной: подсчеты числа хромозом, произведенные во многих случаях разными авторами, показали, что уже при первом делении дочерние ядра получают гаплоидное число хромозом. Правда, Gwynne-Vaughan толкует это таким образом, что здесь редукция выражается не уменьшением числа хромозом, а изменением их валентности (из тетравалентных получаются дивалентные, а из них—моновалентные). Однако такое толкование совершенно произвольно. Иные взгляды защищают Dangeard и некоторые его ученики. Dangeard первый установил кариогамию в сумке (1894) и истолковал это явление как настоящий половой акт (так называемое Данжаровское слияние). Соответственно с этим он отрицает какое бы то ни было слияние в начале развития плодового тела как в смысле цитогамии, так и кариогамии (так называемое Гарперовское слияние). По его мнению, развитие аскогенных гиф идет из одного только аскогона без всякого участия антеридия, у которого он вообще отрицает значение полового органа. Однако при этом игнорируются факты несомненно установленных в некоторых случаях слияния антеридия с ахикарпом. С другой стороны, серьезным аргументом против представлений Данжара является установленная в последнее время раздельнополость гаплоидных мицелиев у некоторых аскомицетов и успешные опыты гибридизации (см. стр. 58). Правда, и против этого выдвигается контрвозражение такого рода, что здесь имеется не настоящая раздельнополость, а только гормональное действие одного мицелия на другой (см. стр. 55). Если это в некоторых случаях так, то, конечно, получение гибридного потомства с анализом передачи признаков того и другого родителя, как это сделано в некоторых случаях, не может быть объяснено только действием гормонов одного мицелия на другой.
Половое воспроизведение, смена генераций и ядерных фаз у низших грибов
У архимицетов для некоторых форм установлена копуляция одинаковых планогамет (подвижных гамет), не отличающихся по существу от зооспор бесполого размножения (некоторые виды Olpidium, Synehytrium endobioticum; рис. 33, 1). В других случаях наблюдается гологамия, т, е. слияние в половом процессе двух вегетативных особей, не диференцированных как специальные гаметы. При этом они или непосредственно прикладываются друг к другу, и содержимое одной (часто меньшей) переливается в другую (Sporophlyctis, некоторые Chytridium), или между ними образуется более или менее длинный копуляционный отросток (Zygorhyzidium, Polyphagia.) Соответственно общему строению Mycochytridiales сливающиеся особи здесь одноядерны, но у Olpidiopsis (из Myxochytridiales), где имеется такое же слияние, они многоядерны и могут быть сравнены с целыми гаметангиями, в которых только не произошло обособления отдельных гамет.
Среди оомицетов наиболее примитивным в смысле полового процесса является Allomyces javanicus, недавно изученный Книпом (1929). Здесь наблюдается анизогамия — слияние двух подвижных гамет неодинаковой величины (♂ и ♀). По строению они (особенно ♀) сходны с зооспорами бесполого размножения, но отличаются от них физиологически (рис. 33, 2). Allomyces и, может быть, другие Blastocladiaceae, которые, кроме названного примера, не изучены в половом отношении, представляют дальнейший этап эволюции изогамных хитридиевых (как Olpidium). У остальных оомицетов имеется резко выраженная оогамия. У Monoblepharis женская гамета — яйцо в виде одноядерной шаровидной клетки свободно лежит в оогонии и даже иногда высовывается из него. Мужские гаметы представлены сперматозоидами — подвижными одножгутиковыми клетками, образующимися по несколько штук в антеридии. Оплодотворение состоит в слиянии сперматозоида с яйцом (рис, 33, 3). У остальных оомицетов (и вообще у всех остальных грибов) обособленных сперматозоидов не образуется; мужской орган представлен здесь антеридием, почти всегда многоядерным и поэтому сравнимым с целым гаметангием, но без диференцировки на отдельные гаметы. Женский орган — оогоний — построен не везде одинаково. У Saprolegniaceae он закладывается как многоядерная клетка, поэтому также сравним с гаметангием. Затем в нем происходит отмирание значительного числа ядер, и протоплазма распадается на некоторое число одноядерных яйцеклеток. Оплодотворение состоит в том, что антеридий пускает отростки, прободающие оболочку рогония (оплодотворяющие отростки); они ветвятся и концами прикладываются к яйцеклеткам, переливая затем в них часть своего содержимого с одним ядром; позднее оно сливается с ядром яйцеклетки (рис. 33, 4). У Peronosporales и некоторых Saprolegniales, например Araiospora, в оогоний диференцируется всегда только одно яйцо, на образование которого идет, однако, не вся протоплазма, а лишь центральная часть ее, периферическая же часть остается в виде так называемой периплазмы и служит для питания яйцеклетки и образования затем толстой оболочки около ооспоры. Оогоний здесь также залагается как многоядерная клетка и может быть сравнен с гаметангием. При диференцировке в нем яйцеклетки эта последняя в большинстве случаев получает только одно ядро, а остальные отходят в периплазму (рис. 33, 5); однако у Albugo bliti и Albugo portulacae яйцеклетка оказывается многоядерной. При оплодотворении у большинства в яйцеклетку переходит только одно мужское ядро, сливающееся с женским, а у Albugo bliti и A. portulacae таким же образом переходит много мужских ядер, которые и сливаются попарно с имеющимися там женскими ядрами.
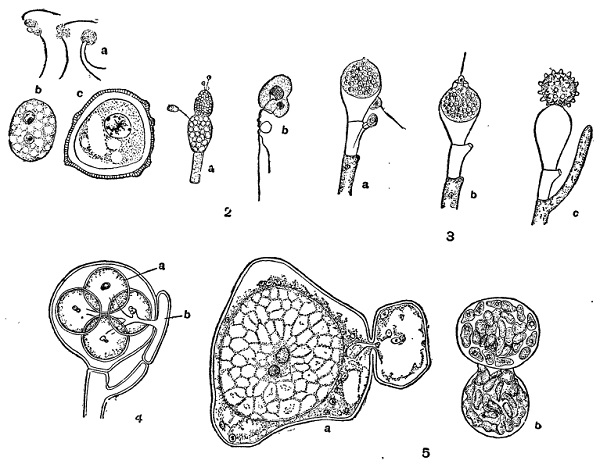
1 — Olpidium viciae:
a — копуляции гамет;
Ь и с — молодая и взрослая зиготы.
2 — Allomyces javanicus:
а — мужской и женский гаметангии;
b — копуляция гамет.
3 — Monoblepharis sphaerica:
а и b — оплодотворение;
с — ооспора.
4 — Saprolegnia:
а — яйцеклетки;
b — антеридий.
б — Albugo Candida:
a — оплодотворение;
b — прорастание ооспоры.
У зигомицетов половой процесс, называемый зигогамией, заключается в слиянии двух большею частью многоядерных клеток — гаметангиев, содержимое которых лишено диференцировки, какая хотя бы имеется в оогониях оомицетов. Они образуются обыкновенно на концах особых коротких ветвей мицелия (копуляционных ветвей) и большей частью совершенно одинаковы по величине и строению (изогамия) (рис. 34, 2). Однако иногда наблюдаются некоторые различия; например у Zygorhynchus или Absidia (из мукоровых) и некоторых других одна клетка всегда больше другой, так что здесь получается некоторый зачаток анизогамии (гетерогамии) (рис. 34, 2). Наконец, у Endogone и Basidiobolus сливающиеся клетки формируются как одноядерные, и в зиготе происходит слияние только одной пары клеточных ядер. Таким образом, и у зигомицетов в эволюции полового процесса происходит замена множественного оплодотворения при слиянии двух гаметангиев простым соединением одноядерных элементов, которые можно сравнить с гаметами, хотя и лишенными диференцировки (рис. 34, 3).
Хотя прорастание половых покоящихся спор у низших грибов (архимицетов, оомицетов и зигомицетов) изучено цитологически очень недостаточно, но все-таки в некоторых случаях установлено, что оплодотворенное диплоидное ядро делится при прорастании редукционным путем (например у Olpidium viciae, — Kusano, 1912; Saprolegnia monoica, — Claussen, 1908; Plasmopara viticola, —Arens, 1929). В других случаях за это говорят генетические данные: разделение полов и расщепление признаков гибридов (у многих мукоровых). Поэтому полагают, что при прорастании споры, происшедшей половым путем, имеет место редукция и возвращение к гаплоидному состоянию.
1 — Rhizopus nigricans;
2 — Absidia spinosa — гетерогамная копуляция;
3 — Endogone — гетерогамная копуляция одноядерных клеток;
4 — Мuсоr—прорастание зигоспоры образованием зародышевого спорангия.
Необходимо отметить, наконец, что покоящиеся споры, сходные с половыми, образуются иногда и без всякого оплодотворения. Такое явление имеет место и среди хитридиевых и особенно известно среди мукоровых и сапролегниевых. У мукоровых иногда развиваются так называемые азиготы — или парой, на месте подошедших друг к другу, но не слившихся гаметангиев, или одиночно, на отдельно стоящей копуляционной ветви. У многих сапролегниевых антеридии образуются далеко непостоянно, а у некоторых видов совершенно отсутствуют; тем не менее в их оогониях яйцеклетки развиваются партеногенетически как бы в настоящие ооспоры (партеноспоры). Возникающие таким путем споры прорастают также после периода покоя совершенно сходным образом с половыми спорами, но, как нужно думать, без редукционного деления ядер.