Специально здесь следует еще отметить некоторые особенности мицелия паразитных грибов, именно тех, которые паразитируют на растениях. По отношению к питающему растению он может быть поверхностным (например у Erysiphaceae) или чаще внутренним, эндофитным. В последнем случае он большею частью идет по межклетникам, но у некоторых распространяется преимущественно в полости клеток (особенно у хитридиевых, где распространение его часто ограничивается всего одной клеткой, но также и у некоторых других форм с более развитым мицелием). Как поверхностный, так и внутренний межклетный мицелий часто развивает особые специальные ветви — гаустории, которые внедряются в полость клетки и воспринимают из нее питательные вещества. Они имеют в разных случаях разную форму, нередко весьма постоянную для отдельных групп. Например у Albugo и Plasmopara гаустории имеют вид как бы маленьких булавочных головок, а у близкого к ним рода, Peronospora, они разрастаются в массивные ветвящиеся образования, заполняющие почти всю клетку питающего растения (рис. 23). Вообще для гаусторий очень типично резкое изменение их характера роста по сравнению с нормальными внеклеточными ветвями мицелия. Это явление вызывается воздействием протоплазмы живой клетки хозяина на внедрившегося нее паразита (подробнее см. стр. 89).
Рис. 23. Гаустории пероноспоровых. 1 — Albugo; 2 — Peronospora parasitica; 3 — то же в поперечном разрезе; видно положение разрезанной гифы в межклетнике и отхождение от нее гаусторий в две клетки.
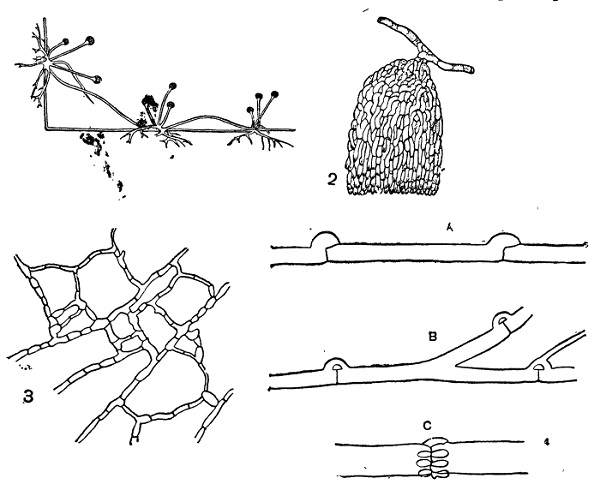
Обнаруживает больше разнообразия. Ветвление гиф обычно обильное, но иногда оно выражено слабо. Например, этим отличаются толстые гифы сапролегниевых и Monoblepharidaceae. Иногда наблюдается известная диференцировка гиф: одни обычно более толстые и слабо разветвленные, а другие — сильно ветвящиеся и более тонкие. Первые служат, главным образом, для распространения мицелия по субстрату, а вторые для воспринятия из него в подходящих местах питательных материалов. Такая диференцировка особенно характерна для мицелия некоторых паразитных грибов, но встречается нередко и среди сапрофитных: например у Rhizopus и некоторых других имеются особые столоны, не ветвящиеся и обладающие энергичным ростом. Они совершенно аналогичны таким же столонами («усам»), например, у земляники и служат также для быстрого вегетативного распространения по субстрату. У таких снабженных столонами форм имеются еще нередко особые органы прикрепления — так называемые ризоиды — в виде пучка коротких разветвленных нитей. Они развиваются как реакция на соприкосновение с твердым субстратом, притом не только с таким, который может иметь известное питательное значение, но и например со стеклом (рис. 22, 1). У некоторых видов Sclerotinia названные органы прикрепления имеют вид довольно массивных кисточек из пучка тесно переплетенных коротких ветвей (рис. 22, 2); то же на неклеточном мицелии наблюдается, например, у Gircinella minor.
Рис. 22. Rhizopus, 1 — столоны со спорангиеносцами на них и ризоидами; 2 — Sclerotinia tuberosa, органы прикрепления мицелия; 3 — анастомозы мицелия; 4 — пряжки: А — развитие пряжки; В — пряжки Merulius; С — Coniophora.
При обильном ветвлении гифы мицелия соприкасаются друг с другом и здесь, на месте соприкосновения, нередко происходит слияние их с установлением сообщения между клетками. Такие слияния называются анастомозами; при большом развитии их мицелий приобретает характерный сетчатый вид (рис. 21 и 22, 3). Развитие анастомозов наблюдается у весьма различных грибов с многоклетным мицелием. В некоторых случаях через них совершается перемещение клеточного ядра из одной клетки в другую, и осуществляется переход от гаплоидного к диплоидному мицелию (у многих гименомицетов,см. стр. 56). Однако большей частью они имеют чисто вегетативное значение и развиваются у многих форм просто при недостатке питания.
Рис. 21. Молодой мицелий шампиньона: 1 — анастомозы, черные — две проросшие базидиоспоры; 2 — начало образования мицелиального тяжа; 3 — клетка, мицелия с окрашенными клеточными ядрами.
В некоторых случаях гифы снабжены еще так называемыми пряжками, представляющими небольшие плоские клетки, лежащие сбоку против поперечной перегородки и сообщающиеся в готовом состоянии открытым отверстием с полостью нижней (проксимальной) клетки. Такие пряжки встречаются только у базидиальных грибов, главным образом у гименомицетов, хотя и не у всех. Их наличие может служить важным признаком для установления принадлежности данного мицелия именно к этой группе. В деталях в свою очередь пряжки бывают различны. Они могут быть или против каждой поперечной перегородки или только против некоторых. Большею частью здесь образуется только одна пряжка, но, например, у очень распространенного разрушителя древесины в постройках, Coniophora cerebella, они развиваются целой мутовкой в количестве до 10 и более штук (этот признак весьма существенен при диагносцирова нии ConiopKora по стерильному мицелию, в каком виде она всего чаще встречается). Наконец, в ряде случаев наблюдается так называемое прорастание пряжек, т. е. развитие из них боковой ветви (например у Merulius и некоторых других, рис. 22, 4b). Пряжки развиваются исключительно на диплоидном (двуядерном) мицелии и служат аппаратом, через посредство которого происходит передача ядра из одной клетки в другую (см. Basidiomycetes, стр. 310). Кроме многих гименомицетов, они встречаются в диплоидном мицелии иногда Ustilaginales и, по некоторым указаниям, также у Uredinales.
Определяется прежде всего наличием в нем воздушных гиф. В одних случаях они идут исключительно по самой поверхности субстрата и отчасти внутри его, так что мицелий получает вид плоского прижатого к субстрату кружка; в других случаях, кроме того, развиваются более или менее обильные гифы, поднимающиеся в воздух и придающие мицелию некоторое сходство, например, с куском ваты, возвышающимся над субстратом. Тот и другой характер роста может быть различным у одного и того же гриба в зависимости от внешних условий, как влажность, питание и т. п., но все-таки для ряда форм имеются специфические особенности: например, образование пышного воздушного мицелия очень типично для многих грибов — разрушителей древесины. Далее, различным может быть цвет мицелия. Чаще всего он снежно-белый, но нередко, особенно с возрастом, приобретает ту или иную окраску, обычно бурую разных оттенков. Это связано с отложением пигмента в клеточных стенках и реже внутри клетки.
Если проследить историю развития какого-нибудь обычного многоклетного мицелия, например у Penicillium glaucum, то можно установить, что мицелий берет свое начало из споры. При прорастании из нее высовывается обыкновенно несколько ростковых трубочек, которые удлиняются и затем отчленяются перегородками от средней части, включающей спору. Они затем еще удлиняются и получают каждый новую перегородку, разделяясь на дистальную, или верхушечную клетку и проксимальную, или внутреннюю. В дальнейшем верхушечная клетка удлиняется и снова делится, отделяя вторую, более молодую по сравнению с первой, внутреннюю клетку. Так повторяется и дальше. Что касается внутренних клеток, то они только вытягиваются, а поперечные деления их наблюдаются редко. Зато из них развиваются боковые ветви. При этом обыкновенно на дистальном конце такой клетки образуется боковая выпуклина, принимающая цилиндрическую форму и отделяющаяся затем перегородкой от производящей ее клетки. Она вырастает затем в боковую ветвь, растущую и ветвящуюся таким же образом, как и главная. Благодаря указанному развитию ветвей на протяжении главной гифы они тем старше и сильнее развиты, чем ближе к основанию лежит место их отхождения (акропетальное ветвление) (рис. 20). Развитие нечленистого (неклеточного) мицелия происходит в общем совершенно такимже образом за исключением, конечно, того, что поперечных перегородок здесь не образуется. Рост сосредоточивается здесь в кончиках гиф, где скопляется обильная протоплазма, заполняющая весь просвет, тогда как в более задних частях получают значительное развитие центральные вакуоли, и протоплазма представлена главным образом тонким стенкоположным слоем. В однородной среде, например на поверхности питательной желатины, гифы мицелия (одинаково как неклеточного, так и многоклетного) разрастаются равномерно-по радиусам во всех направлениях, так что мицелий получает форму круга, нарастающего краями. Центральная часть в нем самая старая и даже иногда отмершая, а периферическая — наиболее молодая.
Рис. 20. Penicillium glaucum, развитие мицелия из споры; А — F — последовательные стадии.
При всем общем однообразии описанного мицелия, который можно назвать типичным или гифенным мицелием, в подробностях, в отдельных случаях, наблюдается ряд специальных черт как макроскопического вида и общего характера роста, так и микроскопического строения.
Протоплазма грибов не обнаруживает в своем строении существенных особенностей. В молодых клетках или в растущих кончиках гиф неклеточного мицелия фикомицетов она заполняет почти сплошь всю полость, однако в ней удается обнаружить путем прижизненной окраски мельчайшие вакуоли то в виде округлых пузырьков (у высших грибов), то в виде узких анастомозирующих друг с другом канальцев (у фикомицетов, например у Saprolegnia, рис. 7,1). В более взрослых частях протоплазма редуцирована до тонкого стенкоположного слоя, а центр клетки занят крупной вакуолей, через которую проходят нередко в разных направлениях тонкие протоплазменные тяжи и пленки (например в крупных клетках сочных плодовых тел гименомицетов).
Рис. 7.1 — Saprolegnia, кончик нити, прижизненно окрашенный «нейтральной красной» краской. Видны вакуоли (υ) в виде анастомозирующих каналов и хондриовомы в виде слегка изогнутых палочек (m); 2 — Leptomitus, часть нити; СС — целлюлиновые тела; N— клеточные ядра, М — хондриозомы.
В узких цилиндрических клетках гиф центральная вакуоля перегораживается обыкновенно поперечными протоплазменными диафрагмами на ряд более мелких, расположенных в ряд по длине клетки.
У некоторых грибов протоплазма обнаруживает движение внутри клеточной оболочки. Особенно это наблюдается у мукоровых с их лишенным перегородок неклеточным мицелием. При этом ток ее направлен преимущественно к верхушечным растущим частям и достигает значительной скорости (до 3 мм в минуту). Часть притекающих таким образом веществ потребляется на месте роста, а вода, повидимому, в значительной массе выдавливается через оболочку наружу. Движение протоплазмы наблюдается иногда в многоклеточном мицелии, например у некоторых дискомицетов. Оно направляется также преимущественно акропетально и идет через не вполне замкнутые поперечные перегородки, пока по середине их имеется еще достаточно широкое отверстие, или пора (рис. 8).
Рис. 8. Гифа Pyronema confluens, движение протоплазмы (стрелки) через поры поперечных перегородок.
Содержащийся в вакуолях клеточный сок, как и в других растительных клетках, представляет в основном водный раствор различных веществ, как, например, Сахаров, органических кислот и их солей, белковых веществ и др. Благодаря своим осмотическим свойствам водного раствора он обусловливает тургор клетки и вместе с тем имеет значение вместилища запасных питательных веществ и продуктов обмена. В клеточном соке могут находиться также оформленные, нерастворимые в воде включения в виде кристаллов, капель масла и др. В огромном большинстве случаев клеточный сок совершенно бесцветен и лишь у немногих представителей содержит растворимые красноватые пигменты, близкие к антоцианам.
Включения в протоплазму
Никаких пластид, не только окрашенных, но и бесцветных лейкопластов грибы никогда не содержат, но в протоплазме их при соответственной обработке обнаруживаются хондриозомы большей частью в виде изогнутых палочек (хондриоконты) (рис. 7 и 9). Согласно главным образом исследованиям Guillermond, много работавшего в этой области, хондриозомы у грибов несут некоторую функцию, аналогичную пластидам у высших растений. Именно в них отлагаются сначала тельца метахроматина, затем освобождающиеся в протоплазму и из нее переходящие в вакуоли. Метахроматин, называемый также волютином, представляет очень характерный запасный продукт в клетке грибов (также бактерий и некоторых водорослей) и имеет, судя по всему, протеиновый характер, представляя соединение нуклеиновой кислоты с каким-то органическим основанием. Он окрашивается ядерными красками, но в несколько иной тон (метахромазия) и особенно характеризуется прижизненной окраской от метиленовой сини (рис. 9).
Рис. 9. Строение клетки в мицелии Sclerotinia libertiana. 1—-3 живые клетки; 4—6 фиксированные и окрашенные. 1 — правая клетка молодая (вторая от вершины гифы) — протоплазма с очень мелкими вакуолями; левая клетка—более взрослая с более крупными вакуолями в протоплазме; 2 — более взрослая стадия; вакуоли еще крупнее; 3 — еще более взрослая клетка, обильные крупные вакуоли придают протоплазме пенистый вид; 4 — окрашенные ядра в клетке, соответствующей по возрасту рис. 2; 5 — окраска хондриозом в клетке того же возраста; 6 — одновременная окраска ядер и хондриозом в клетке того же возраста.
Из других запасных продуктов в клетках грибов широко распространены капельки жира и гликоген. Они образуются в протоплазме, но могут затем переходить из нее в клеточный сок.
Жировые вещества, или липоиды, нередко скопляются в протоплазме в очень мелко распыленном состоянии, как бы пропитывая ее и придавая ей при этом более блестящий стекловатый вид. В дальнейшем эти вещества могут сливаться в более крупные уже различаемые под микроскопом блестящие капельки, которые или остаются в протоплазме, или выпадают из нее в вакуоли.
Такой процесс обособления жировых капелек ускоряется при некоторых неблагоприятных воздействиях на клетку (например при изменении реакции среды) вследствие чего такие пострадавшие подавленные клетки обнаруживают особенно большое количество жировых капелек в вакуолях, а первоначальная конфигурация протоплазмы в них при отдаче значительного количества липоидов нарушается. Такое явление носит специальное название липофанероза. И без специальных экспериментов его легко наблюдать в старых культурах грибов или в грибах, находящихся в неблагоприятных условиях среды. В других случаях жир с самого начала отлагается в виде более крупных капелек, не переходя через мелко распыленное состояние.
Также и гликоген отлагается в протоплазме в мелко раздробленном состоянии коллоидного раствора. Пропитывая известные участки протоплазмы, например в молодых сумках, он придает им блестящий стекловатый вид, напоминающий отчасти скопление липоидов. Однако присутствие гликогена легко обнаруживается при действии раствором йода, дающего с ним красно-бурое окрашивание. Попадая в вакуоли, гликоген и в них остается обыкновенно в виде коллоидного раствора и обнаруживается действием йода.
Из других оформленных включений в клетке можно указать на белковые кристаллы, встречающиеся, например, часто у Мисогасеае и некоторых других, преимущественно низших грибов. Они могут находиться как в протоплазме, так и в вакуолях.
Далее можно отметить целлюлиновые тела (у Leptomitus, рис. 7, 2) и фиброзиновые тела (в конидиях Erysiphaceae). Состав их неизвестен. Фиброзиновые тела играют роль запасного продукта, так как при прорастании конидии они исчезают.
В клетках плодовых тел многих трутовиков и других часто наблюдается отложение смолы в виде бесформенных бурых телец. Смола здесь — продукт отброса. В мелко раздробленном состоянии она содержится также в млечных гифах видов Lactarius й других. Кристаллы оксалата кальция, столь обычные на поверхности клеточной оболочки, внутри клетки встречаются крайне редко (малое поступление Са внутрь клетки). Они известны, например, в клетках плодового тела Russula и особенно в мицелии Mutinus. У последнего они образуют нечто вроде крупного сферокристалла, растягивающего клетку (рис. 6, 3).
Рис. 6. 1 — Merulius, гифы из тяжа, Н — обыкновенные вегетативные гифы; F — механические гифы с утолщенными стенками, G — сосудистые гифы с неравномерно утолщенными стенками; 2 — Battarea, клетка капиллиция; 3 — Mutinus caninus, гифа с сферокристаллом оксалата внутри и отдельными кристаллами на поверхности.
Клетка грибов почти всегда одета твердой оболочкой; исключением являются зооспоры, а у миксохитридиевых — также и вегетативные состояния, представленные голой протоплазмой.
Оболочка на молодых клетках обыкновенно тонкая, бесцветная и бесструктурная и остается иногда такой до конца (особенно в мицелии). В других случаях наблюдаются часто дальнейшие ее изменения в виде утолщения, появления слоистости, отложения пигментов и инкрустаций и т. д., благодаря чему оболочка может получать иногда довольно сложную структуру (особенно в спорах и в меньшей степени в вегетативных клетках).
Утолщение оболочек вегетативных клеток происходит изнутри и является почти всегда сплошным. При значительном развитии оно может вести к заметному сужению и даже местами к облитерации клеточной полости. Такие утолщения наблюдаются в частях, отличающихся особой прочностью, например в твердых деревянистых плодовых телах некоторых трутовиков, в мицедиальняых тяжах домового гриба (МегиНиз lacrymans), а также в клетках-ножках некоторых спор, например телейтоспор некоторых ржавчинных грибов и др. Лишь в виде исключения наблюдаются внутренние местные утолщения, например, иногда в гифах сапролегниевых или «сосудистых гифах» тяжей домового гриба — в виде неправильных внутренних бугров и бородавок, или в капиллиции Battarea — в виде довольно правильных колец и спиралей (рис. 6, 1 и 2).
Рис. 6. 1 — Merulius, гифы из тяжа, Н — обыкновенные вегетативные гифы; F — механические гифы с утолщенными стенками, G — сосудистые гифы с неравномерно утолщенными стенками; 2 — Battarea, клетка капиллиция; 3 — Mutinus caninus, гифа с сферокристаллом оксалата внутри и отдельными кристаллами на поверхности.
В утолщенной оболочке нередко можно различить наружный (первичный) слой и внутренний (слой утолщения). Часто они заметны только при действии веществ, вызывающих разбухание (серная кислота и др.); при этом выявляется большая стойкость наружного слоя, где нередко имеется кутинизация. Первоначально бесцветная оболочка с возрастом часто пигментируется большей частью в бурые, иногда почти черные тона, реже в яркие, как голубой у Barlaea (Plicariella) fulgent, зеленый у Chlorosplenium aeruginosum и др. (см. пигменты грибов). Кроме настоящего утолщения оболочки, происходящего изнутри, у грибов нередко наблюдается ослизнение и разбухание в воде наружных частей ее. Это явление очень широко распространено и для некоторых групп особенно типично (например у Tremellales). При сближении таких ослизненных гиф их слизистые чехлы склеиваются часто до полной потери собственных контуров, так что, например, микроскопическое исследование в воде плодовых тел Tremella показывает гифы, одетые твердыми (внутренними) оболочками, рыхло проходящие в общей массе бесструктурной слизи. Последняя иногда окрашивается пигментами, например красным в верхних слоях шляпки мухомора (Amanita muscaria) или желтым у Boletus lute us.
По химической природе грибные слизи представляют преимущественно производные маннозы.
Состав клеточной оболочки.
Что касается состава твердой клеточной оболочки, то в основе своей она сложена из углеводов. При этом у оомицетов она обыкновенно дает посинение от хлорцинкиода и должна рассматриваться как целлюлоза. Иногда к ней (у пероноспоровых) примешивается в значительном количестве каллеза.
Что же касается мукоровых и всех высших грибов (аскомицетов и базидиомицетов), то клеточные оболочки у них имеют, повидимому, в основе какое-то видоизменение целлюлозы, называемое иногда метацеллюлозой. Особенно же характерно, что к этой углеводной основе присоединены еще азотистые вещества, сходные с хитином насекомых.
Следует отметить, что у многих сумчатых грибов оболочки сумок синеют от йода подобно крахмалу. Предполагается, что они состоят из углеводов, близких к изолихинину, изомерному с крахмалом и более определенно изученному у некоторых лишайников. Благодаря этому клеточная оболочка высших грибов не растворяется в реактиве Швейцера, не дает реакции с хлорцинкйодом и т. д. Как у высших, так и у многих низших грибов оболочка нередко еще кутинизирована, а в спорах, кроме того, пропитана еще жиром или воском, благодаря чему плохо смачивается водой. Лигнина и суберина в оболочках грибов не было обнаружено с достаточной убедительностью, хотя плодовые тела некоторых трутовиков весьма напоминают по своей консистенции древесину или пробку (у Daedalea quercina, по некоторым указаниям, имеется опробковение оболочек).
Оболочка нередко инкрустируется оксалатом кальция, или в виде отдельных неправильной формы кристалликов (в мицелии очень различных грибов), или в виде тесного слоя игольчатых кристаллов (рис. 1, 2 и рис 6, 3).
Рис. 1. Mucor mucedo. 1 — спорангиеносцы со спорангиями и мицелий; 2 — спорангий, сильно увеличенный; 3 — зигоспора на месте слияния двух одинаковых ветвей мицелия.
Хотя у грибов очень нередко развиваются весьма массивные органы со сложным внутренним строением, как плодовые тела, склероции и т. п., однако при анатомическом изучении большей частью оказывается, что они сложены не из настоящих тканей, которые происходили бы в результате деления клеток во всех направлениях, а из так называемых ложных тканей. Настоящие ткани в анатомическом смысле встречаются у грибов редко. Из них слагается, например, тело (receptaculum) у Laboulbeniales, и кое-где они встречаются у отдельных представителей других групп. Например, таким образом проходят ранние стадии развития перитеция у Sporormia intermedia, то же наблюдается при развитии некоторых пикнид, например у Pleospora herbarum и др. Вообще пикниды, повидимому, образуются нередко в результате деления клеток во всех направлениях. В огромном большинстве случаев различные более массивные тела грибов образованы в результате сплетения и срастания нитчатых элементов — гиф, из которых каждая имеет деления только в одном поперечном направлении. Такие соединения на зываются ложными тканями, или плектенхимой (также псевдопаренхимой) . Плектенхима бывает то более плотной, так что между клетками остаются только маленькие промежутки, наполненные воздухом, то более рыхлой. В одних случаях клетки слагающих ее гиф делятся на более короткие отрезки, которые — вздуваются и округляются, так что на разрезах получается впечатление ткани, сложенной из более или менее изодиаметрических клеток, т. е. паренхимы. Согласно ее происхождению такую ткань называют параплектенхимой. В других случаях клетки сохраняют свою нормальную для гиф вытянутую цилиндрическую форму, и на разрезах получается ткань, сложенная из удлиненных клеток, идущих то параллельно друг другу, то переплетаясь. Такие грибные ткани называют прозоплектенхимой. В отличие от параплектенхимы ее сложение из нитчатых элементов выступает с большей ясностью (рис. 16, 1).
Хотя, как было указано выше, настоящих тканей у грибов почти не встречается, но тем не менее с точки зрения физиологической анатомии можно отметить различные гифы и комплексы их, приспособленные к выполнению определенных функций. Их можно трактовать как физиологические ткани и разделить на такие же группы, как и ткани высших растений, т. е. меристема, кроющие, механические, проводящие ткани и т. д. Однако эти категории здесь гораздо менее диференцированы, нередко связаны друг с другом переходами и мало обособлены в пространстве.
Настоящая меристема у грибов не развивается. Деление клеток сосредоточено на концах гиф. Однако в некоторых случаях эти делящиеся участки соединяются в тесную параплектенхиму, весьма напоминающую под микроскопом меристему. Наилучший пример такого рода можно видеть на кончике ризоморфы опенка, весьма похожем анатомически на кончик корня (рис. 16, 2). В плодовых телах Gorticium и др., нарастающих неопределенно своими краями, последние также могут быть сравниваемы с меристемой. Однако здесь они имеют ясное гифенное строение.
Кроющая ткань
У грибов развита особенно на поверхности покоящихся органов, как склероции, а также на верхней поверхности шляпки многих гименомицетов. Она имеет иногда характер однослойной или многослойной параплектенхимы, причем ее клетки имеют утолщенные и нередко окрашенные оболочки. У склероциев Sclerotinia fuckeliana кроющая ткань имеет вид однослойного эпидермиса, а у склероциев Typhula это сходство с эпидермисом увеличивается еще сильными утолщениями наружных стенок и тем, что при рассматривании с поверхности они имеют извитые контуры подобно эпидермису на листьях двудольных растений (рис. 17, 1). У Polyporus lucidus кроющая ткань образована слегка расширенными концами гиф, ясно продолжающихся во внутреннюю плектенхиму. Они расположены одним палисадным слоем, имеют утолщенные оболочки и тесно склеены друг с другом слизью, благодаря чему поверхность плодового тела гриба получает характерный лакированный вид (рис. 17, 2).
Рис. 17. 1 Typhula gyrans, наружный слой склероция с поверхности; 2 — Polyporus lucidus, наружные слой плодового тела в разрезе.
Во многих других случаях кроющая ткань плодовых тел более массивна, многослойна и состоит из переплетающихся друг с другом гиф (т. е. имеет характер прозоплектенхимы). Например, у масленка слагающие ее гифы расположены не плотно, но промежутки между ними заполнены густой упругой слизью, так что в результате получается довольно прочная пленка, которая резко отличается по цвету и по косистенции от нижележащей ткани и легко сдирается с нее. Такое же строение кроющей ткани набтюдается у многих других шляпных грибов (особенно тех, у которых верхняя поверхность шляпки липкая).
Приведенные примеры представляют наиболее диференцированные типы кроющих тканей. Во многих других случаях они выражены гораздо слабее, не резко отграничены от внутренних тканей и с наружной стороны кончаются не гладкой поверхностью, а или массой волосков из засохших гиф (на ризоморфах опенка, плодовых телах многих Thelephoraceae), или все время отпадающими округлыми клетками, благодаря чему поверхность получает вид как бы осыпанной мукой (плодовые тела многих Lycoperdaceae).
Наконец, во многих случаях плодовые тела и совсем лишены какой бы то ни было диференцированной кроющей ткани.Так это, например, обстоит у шляпных грибов в нижней стороне шляпок, нередко на пеньках, также на плодовых телах Clavariaceae и др. Вопрос о физиологическом значении кроющей ткани у грибов, даже в случав высокой степени ее морфологической диференцировки, не может считаться выясненным. Специальных исследований в этом направлении, насколько известно, не производилось. Согласно нашим опытам с масленком (Boletus granulatus) сдирание пленки с верхней стороны шляпки не повышает заметно ее испарения по сравнению с неободранной как в первые часы опыта, так в течение суток, когда потеря воды достигает уже 50% от первоначального веса. Таким образом, в данном случае не приходится говорить о защите от испарения; вероятнее видеть здесь функцию поглощения воды разбухающей слизью. Такое же значение должна иметь волосистая кроющая ткань, например на плодовых телах Polystictus versicolor и др., быстро высыхающих и при увлажнении снова быстро набухающих. Возможно, что в других случаях кроющие ткани несут функции защиты от испарения или механических воздействий.
Механические ткани
Представлены гифами с утолщенными стенками и соответственно суженным просветом. Они широко распространены особенно в твердых плодовых телах трутовиков и некоторых мицелиальных тяжах, например у Merulius lacrymans, но большей частью мало отграничены и перемешаны с другими элементами (рис. 6,1). В некоторых случаях, например в ножках апотециев Sclerotinia, наблюдается некоторое оттеснение утолщенных элементов на периферию, что, конечно, сообщает ножке большую прочность на изгиб. В большинстве случаев известная прочность на изгиб прямостоящих частей сочных плодовых тел достигается просто взаимным напряжением тканей, растянутых в наружных частях и сжатых во внутренних, подобно тому как это имеет обыкновенно место в сочных травянистых стеблях цветковых растений.
Рис. 6. 1 — Merulius, гифы из тяжа, Н — обыкновенные вегетативные гифы; F — механические гифы с утолщенными стенками, G — сосудистые гифы с неравномерно утолщенными стенками; 2 — Battarea, клетка капиллиция; 3 — Mutinus caninus, гифа с сферокристаллом оксалата внутри и отдельными кристаллами на поверхности.
Проводящие ткани
Большинство грибов даже с крупными сложно построенными плодовыми телами не имеет особо диференцированных элементов для передвижения воды, и это, между прочим, ограничивает распространение их более или менее сырыми местообитаниями, где испарение и стало быть потребность в проведении воды меньше. Передвижение веществ происходит здесь по обыкновенным гифам и облегчается тем, что поперечные перегородки в них, как было указано выше, снабжены отверстием. Однако во многих случаях у грибов отмечаются более совершенные проводящие элементы, напоминающие даже сосуды. В типичном случае они представлены широкопросветными прямыми маловетвящимися гифами нередко с неравномерно утолщенной боковой стенкой (встречаются утолщения в виде неправильных колец и т. п.) и частичным или даже полным растворением поперечных перегородок между клетками. Живое содержимое их в значительной степени редуцировано, и они наполнены главным образом водянистой жидкостью. Такие водоносные элементы идут, не соединяясь в пучки, среди других тканей. Наилучшего развития они достигают в мицелиальных тяжах домового гриба (Merulius lacrymans) (рис. 6, 1).
Более распространены проводящие элементы для органических веществ, встречающиеся преимущественно в плодовых телах некоторых гименомицетов. Они представлены также изолированно идущими гифами, отличающимися более густым, нередко окрашенным содержимым, богатым питательными продуктами. Такие элементы залагаются очень рано в плодовом теле в качестве ответвлений обычных гиф, но идут далее самостоятельно, иногда анастомозируя друг с другом. Поперечные перегородки в них отсутствуют на значительных протяжениях. Содержимое представлено протоплазмой с многочисленными клеточными ядрами и клеточным соком указанного выше состава. Физиологическое значение этих гиф подчеркивается тем, что они особенно близко подходят к спороносным частям плодового тела, где идет усиленное потребление питательных материалов (рис. 18, 1 и 2).
Рис. 18. 1 — Corticium violaceo-lividum, разрез через край плодового тела, пронизанного проводящими гифами; 2— Lactarius deliciosus (рыжик), прохождение млечных гиф в пластике; конечные ответвления их проникают между базидиями.
В некоторых случаях наблюдается образование продырявленных ситовидных пластинок между этими гифами и окружающими их клетками (например у Mycena galeopus). Иногда содержимое таких гиф ярко окрашено растворенным пигментом (например у Fistulina hepatica), в других случаях оно имеет вид тонкой эмульсии от присутствия мелких капель распыленной смолы или жира, бесцветных или окрашенных. Такая масса называется млечным соком, а содержащие ее гифы млечными трубками (например у рыжика и др., рис. 19).
Рис. 19. Млечные трубки (т) у Lactarius, проходящие преимущественно через псевдопаренхиматическую ткань: 1 — продольный разрез пенька; 2— поперечный разрез пенька.
По содержанию запасных питательных веществ и расположению в плодовом теле они относятся к тем же проводящим элементам, а по содержанию смолы — к вместилищам экскретов. У многих гименомицетов, особенно у трутовиков, в плодовых телах содержатся еще более или менее округлые клетки, заполненные смолистым содержанием. Их можно рассматривать как типичные вместилища выделений.
У грибов встречаются как одноядерные клетки, так и многоядерные. Низшие грибы, как мукоровые, сапролегниевые и другие с сильно развитым неклеточным мицелием, содержат в нем очень большое количество клеточных ядер (рис. 10).
Рис. 10. Saprolegnia. Окраска ядер. 1—растущий кончик гифы; 2 — средняя часть той же гифы.
Наоборот, хитридиевые большей частью одноядерны. Строго одноядерны среди аскомицетов клетки у всех Erysiphaceae (рис.11, 7). То же изолированно встречается у представителей других групп. Однако гораздо чаще среди аскомицетов встречаются формы с многоядерными клетками. Что касается базидиомицетов, то здесь обыкновенно больше определенности в числе клеточных ядер. Как правило мы имеем здесь или одноядерные или двуядерные клетки, причем те и другие характерны для разных стадий одного и того же гриба (см. смена ядерных фаз) (рис. 11, 2—4), Впрочем, у базидиомицетов первоначальная одноядерность (или двуядерность)в более старых клетках нередко сменяется на вторичную многоядерность.
Рис. 11. 1 — Phyllactinia, клетки мицелия с гаусторией, все одноядерны; 2 — Uromyces scutellatus, одноядерные клетки мицелия; 3 — Corticium terrestre, мицелий из двух двуядерных клеток; 4 — Corticium terrestre, одновременное деление двух ядер двуядерной клетки (дикариона).
Размеры клеточных ядер у грибов большею частью весьма незначительны (2—3 µ в диаметре). Наиболее крупные ядра встречаются в органах, связанных с половым воспроизведением, например в базидиях и особенно сумках, где первичное копуляционное ядро нередко достигает 10µ и даже более в поперечнике; сравнительно крупные ядра (и в вегетативных органах) имеет большинство Uredinales (рис. 12, 1). Наоборот, Мисогасеае и Ustilaginales отличаются чрезвычайной мелкостью своих клеточных ядер (в среднем около 1—1,5 µ в диаметре) (рис. 12, 2).
Рис. 12. 1 — Puccinia helianthi двуядерные клетки (увел. 750 раз); 2 — клеточные ядра в мицелии Мисог (увелич. 1500 раз); 3 — митозы в сумке Galactinia succosa; 4 — прямое деление ядра в почкующейся клетке дрожжей.
По форме ядра большею частью округлы или слегка вытянуты вдоль клетки. Строение их, поскольку его удалось изучить у сравнительно крупноядерных форм, представляет следующую картину: более или менее густая хроматиновая сеть и обычно одно ядрышко, лежащее часто весьма эксцентрично и даже иногда как бы высовывающееся отчасти за пределы ядерной оболочки. Последняя не всегда ясно выражена (рис. 11, 2; 12, 1).
Что касается более мелких ядер, то строение их разобрать очень трудно. Часто они представлены в виде мельчайшего окрашивающегося тельца, окруженного светльм, лишенным структуры, двориком (рис. 12, 2). Это тельце, лежащее большею частью центрально, обычно называется ядрышком, но не исключено, что в некоторых случаях оно представляет центральное скопление хроматина (т. е. ядро здесь кариозомного типа).
Деление ядер изучено главным образом в репродуктивных органах, особенно в развивающихся сумках и базидиях, где оно носит редукционный характер. В сумках наблюдается типичный мейозис с хорошо выраженными хромозомами, ахроматиновым веретеном и нередко с центрозомами на его полюсах и лучистыми фигурами, расходящимися от них в протоплазме. Фигура мейозиса формируется внутри ядерной оболочки, которая здесь вообще сохраняется обыкновенно очень долго, исчезая нередко только в телофазе (рис. 13).
Рис. 13. Развитие сумки у Peziza. 1 — слияние гаплоидных ядер; 3—8-стадии деления копуляционного ядра; 9 — второе деление; 10 — третье деление.
В базидиях имеют место такие же хорошо сформированные мейозы с тем отличием, что ядерная оболочка исчезает здесь большей частью уже во время профазы. Такие же вполне четкие митозы наблюдаются в развивающихся оогониях сапролегниевых и пероноспоровых. Что касается вегетативной области, то здесь деление ядер изучено крайне слабо. Оно нередко здесь носит примитивный, или упрощенный характер: ахроматиновое веретено деления выражено плохо, и самые хромозомы не обособляются с ясностью друг от друга. Однако при их расхождении в телофазе вырисовываются и долго сохраняются затем соединительные нити. На этой стадии хромозомы часто соединяются в две группы на каждом полюсе (рис. 14, 5). Это и дало основание ряду преимущественно французских авторов прийти к заключению, что число хромозом у всех базидиомицетов очень постоянно и равно двум а у всех аскомицетов оно равняется четырем. Такое заключение, поддерживаемое кое-где и в современных французских работах, нужно признать ошибочным. Те красящиеся тельца, которые видны в телофазе вегетативного деления, нужно толковать не как отдельные хромозомы, а как результат их соединения, так сказать, как метахромозомы. Это следует, например, из изучения кариокинеза у Uredinales. В вегетативной области у них обычно можно наблюдать по паре хроматиновых телец на каждом полюсе телофазы, а в редукционном делении, где хорошо обособляются как ахроматиновое веретено, так и хромозомы, число последних оказывается значительно больше двух: у разных видов от 8 до 16 (рис. 14).
Также и у аскомицетов число хромозом в редукционном делении, где их легче можно сосчитать, оказывается различным у разных видов, и почти всегда значительно больше четырех (например у Pyronema гаплоидное число их равно 12). Еще более упрощенные деления ядер в вегетативной области указываются у Entomophthoraeeae и дрожжей. Здесь ядерная оболочка сохраняется до конца деления, ахроматиновые нити не выражены, и ядро как бы просто перешнуровывается. Однако внутри его в это время происходят некоторые перегруппировки хроматина, как бы попытки к обособлению хромозом. Такое деление в известной мере напоминает амитоз. Еще более типичные амитозы описаны в старых уже дегенерирующих частях, например в колонках спорангиев мукоровых. Однако у сапролегниевых, по описанию Smith (1923), амитотически делятся молодые ядра в растущих кончиках гиф. Интересно, что у них же в развивающихся затем оогониях имеются вполне типичные митозы с ясно обособленными хромозомами. Таким образом, здесь имеется необычное цитологическое явление, что ядра, делившиеся сначала амитотически, восстанавливают затем в известный период типичные митозы. Обычно полагают, что амитоз имеет место только в дегенерирующих клетках, и митоз в них уже не восстанавливается. Довольно вероятно, амитотические деления или только внешние, похожие на них, распространены в вегетативной области и у многих других грибов.
Интересно отметить еще характер ядерных делений в тех клетках базидиальных грибов и аскогенных гифах сумчатых, которые нормально являются двуядерными. Их ядерный аппарат, называемый дикарионом, или синкарионом, состоит из двух ядер, то сближенных друг с другом, то отдаленных. При наступлении деления ясно выступает динамическая связь между ними, выражающаяся в том, что оба ядра делятся строго одновременно. При этом, если, как это часто бывает, ядра в покоящемся состоянии лежат друг за другом по длине клетки, иногда даже в ее противоположных концах, то, приступая к делению, они сближаются, ложатся рядом в поперечном направлении и затем одновременно делятся, причем оси делений ориентируются обыкновенно вдоль клетки (рис. 11, 4). Когда образовавшиеся две пары ядер разойдутся и клетка разделится поперек на две, то каждая из них в результате указанного механизма получит по два ядра, не являющихся сестрами.
Деление клетки у грибов не столь тесно связано с делением ядра, как это наблюдается у большинства других растений. Это имеет место не только в многоядерных клетках, но и в нормально одноядерных. Деление клетки может значительно отставать от деления ядра, благодаря чему на протяжении мицелия, состоящего из одноядерных клеток, могут нередко попадаться двуядерные (еще не успевшие разделиться) клетки. Цитологической основой указанного явления, может быть, является то, что, насколько это известно, при делении ядер у грибов никогда не образуется фрагмопластов, и самая клетка делится потом независимо от ядра путем врастания перегородки от периферии к центру. Эта перешнуровывающая клетку поперечная перегородка в самом центре часто не смыкается окончательно, оставляя здесь узкое отверстие (рис. 15). Благодаря этому клетки многоклеточного мицелия не вполне изолируются друг от друга, а остаются соединенными тончайшими протоплазменными нитями, называемыми плазмодесмами. В мицелии эти отверстия обыкновенно крайне узки (поА. Меуеr’у около 0,3 µ в диаметре), однако иногда они бывают и шире. Это наблюдается, например, при развитии некоторых конидий, расположенных цепочками. При этом через расширенные отверстия (до l µ и шире) легче осуществляется передача питательных веществ от нижних клеток к верхним. Через такие отверстия в поперечной стенке может иногда переходить даже клеточное ядро из одной клетки в соседнюю. Повидимому, такое явление нередко имеет место как у сумчатых, так и у базидиальных грибов в процессе апогамии (см. стр. 43). Такие же отверстия в перегородках образуются при нередко наблюдаемых у грибов анастомозах между соседними гифами. Эти вторичные отверстия обыкновенно значительно шире. Через них также может происходить переход ядер.
Рис. 15. Поры и плазмодесмы. 1 — базидия у Coprinus; 2—конидиеносец у Penicillium; 3 — гифа у Merulius с поперечной перегородкой и пряжкой. Оболочки взбухли от действия серной кислоты.
Грибы представляют собой очень обширную группу (тип) растений, содержащую до 80 000 описанных видов. Возможно, впрочем, что некоторое количество из указанного числа представляет одни и те же виды, но повторно описанные под разными названиями. Однако, с другой стороны, нужно признать, что видовой состав грибов изучен еще далеко не полно. Особенно это относится к тропическим областям, где, вероятно, будет найдено еще огромное количество новых видов. Поэтому некоторыми микологами-систематиками и флористами высказывается мнение, что грибы по общему числу видов, существующих сейчас на земле, мало чем уступают, а может быть, даже превосходят цветковые растения (около 150 000 видов). При указанном богатстве видами грибы отличаются вместе с тем большим разнообразием их. Стоит только вспомнить, что сюда относятся столь различные представители, как всем известные шляпные почвенные грибы, или древесные трутовики с их крупными сложно построенными плодовыми телами, далее — крупные, иногда огромные дождевики (до 1,5 м в диаметре у американской Calvatia) затем различные подземные клубневидные грибы, как трюфели и др. С другой стороны, к грибам относится еще значительно большее число микроскопических форм, заметных невооруженному глазу в виде различных налетов и плесеней на тех или других субстратах, а нередко и совсем неразличимых без микроскопа.
При всем этом разнообразии можно указать ряд черт, свойственных всем или огромному большинству грибов. Во-первых, все без исключения грибы не содержат хлорофилла и поэтому имеют гетеротрофное питание, т. е. черпают свой углерод только из готовых органических соединений. В соответствии с этим стоит строение их вегетативного тела. За исключением небольшого числа наиболее примитивных видов, живущих как внутриклетные паразиты зеленых растений, почти у всех остальных оно развито в виде мицелия, или грибницы, и представляет систему очень тонких ветвящихся нитей, или гиф, непрерывно нарастающих своими концами и пронизывающих субстрат, на котором развивается гриб. Благодаря такому строению мицелия, он имеет очень большую поверхность, через которую осмотическим путем и поступают в него питательные вещества, в том числе и сравнительно мало подвижные, но необходимые для него органические вещества.
При микроскопическом изучении обнаруживается, что мицелий различно построен у низших и высших грибов. У первых слагающие его гифы, хотя и достигают многих сантиметров в длину, но лишены поперечных перегородок, и, таким образом, весь мицелий оказывается, формально говоря, одной сильно разветвленной клеткой с огромным числом ядер в ней. Такой мицелий называется одноклеточным, или не членистым. У всех высших грибов и немногих низших гифы мицелия перегорожены поперечными перегородками на отдельные клетки, каждая с одним или чаще с несколькими ядрами. Такой мицелий называется многоклеточным, или членистым. За этим, хотя и существенным различием, мицелий всех грибов построен весьма сходно и одинаково пронизывает субстрат или (при большой влажности атмосферы) отчасти растет сверх него, образуя пушистый налет или более объемистые ватообразные скопления большею частью беловатого цвета.
Мицелий построен в общем слишком однообразно, чтобы по нему можно было идентифицировать тот или другой гриб. Более разнообразны и характерны для того или иного вида или группы видов органы размножения, приносящие споры и называемые поэтому также органами спороношения. При этом здесь различают бесполые и половые спороношения. Первые, подобно вегетативному размножению высших растений, служат для быстрого размножения и повторяют, как правило, полностью особенности произведшего их организма. Вторые имеют в основе половой процесс, сопровождающийся слиянием клеточных ядер и следующим затем редукционным делением их. Поэтому споры полового размножения, подобно семенам высших растений, не являются все вполне тождественными генетически друг с другом и с материнским растением.
Половые споры нередко представляют покоящиеся споры и служат в этом случае не столько для непосредственного размножения, сколько для сохранения вида в течение неблагоприятного периода (зима, засуха).
Среди бесполых спор различают эндогенные и экзогенные. Первые имеются только у низших грибов и образуются в большом числе внутри особых клеток, называемых спорангиями, почему и самые споры называют спорангиоспорами. Экзогенные споры, иначе называемые конидиями, свойственны высшим грибам и только немногим низшим. Они образуются не внутри, а на поверхности образующего их органа, называемого конидиеносцем.
Что касается половых спор, то они также бывают различны. У низших грибов они имеют характер покоящихся спор и различаются по форме полового процесса, давшего им начало (ооспоры, зигоспоры). При прорастании их развивается спорангий со спорами в нем, происшедшими в результате редукционного деления (т. е. генетически не однородными). У высших грибов половой цикл сложнее. Продукт оплодотворения у них не превращается в покоящуюся спору, а разрастается далее и в конце своего развития приносит многочисленные спороносные органы. Одни из них образуют внутри эндогенные споры и называются сумками, или асками, а их споры — аскоспорами. Другие образуют на своей поверхности экзогенные споры и называются базидиями, а их споры — базидиоспорами. Как аскоспоры, так и базидиоспоры образуются в результате редукционного деления, т. е. являются генетически неоднородными, как это свойственно вообще спорам полового происхождения (см. ниже споры спорангиев, получающихся при прорастании половых спор низших грибов).
Для пояснения вышеизложенного можно рассмотреть несколько конкретных примеров.
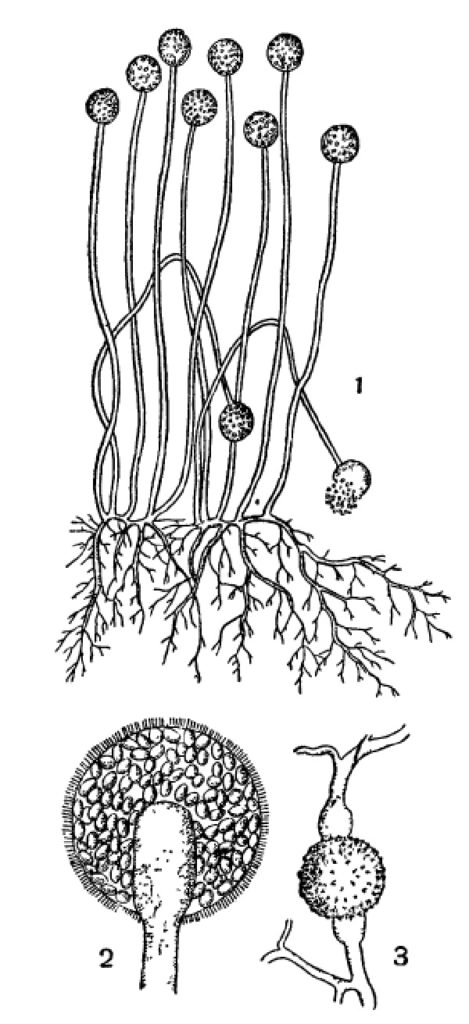
Обычнейший плесеневый гриб, Mucor mucedo, принадлежит к низшим грибам. Его бесполые спорангиоспоры выпадают из спорангия и, перенесенные токами воздуха на субстрат подходящего состава и влажности, немедленно прорастают. При этом спора вздувается и затем из нее вырастают 2—3 ростковые трубочки, которые быстро растут своими концами дальше, ветвятся и развиваются в нечленистый (одноклеточный или, лучше, неклеточный) мицелий (рис. 1). Он отчасти пронизывает субстрат, отчасти стелется по его поверхности. Через 2—3 дня роста на таком мицелии развиваются особые ветви, которые не ветвятся и растут прямо вверх на высоту до 10 см. Они представляют спорангиеносцы. Их концы шаровидно вздуваются, отделяются от спорангиеносца перегородкой и превращаются в спорангии (рис. 1). В спорангиях без всякого редукционного деления образуются спорангиоспоры таким образом, что общее протоплазменное содержимое распадается на большое число участков, одевающихся каждый собственной оболочкой (рис. 1, 2). Спорангиоспоры, выпадая вследствие разрушения оболочки спорангия, немедленно могут прорастать в такой же мицелий с новыми спорангиями на нем и т. д.
Рис. 1. Mucor mucedo. 1 — спорангиеносцы со спорангиями и мицелий; 2 — спорангий, сильно увеличенный; 3 — зигоспора на месте слияния двух одинаковых ветвей мицелия.
У Mucor mucedo имеется и половой процесс. Он происходит лишь при определенных условиях: при встрече двух различных в половом отношении мицелиев. При этом специальные короткие ветви гиф того и другого сливаются «своими концами, и из этого слияния получается зигоспора, одевающаяся толстыми оболочками и переходящая в состояние покоя (рис. 1,3). Позднее она прорастает в короткую нить с спорангием на конце. В нем на этот раз происходит редукционное деление при формировании спорангиоспор, так что последние в этом так называемом зародышевом спорангии оказываются генетически разнородными, отличаясь, между прочим, половыми свойствами развивающихся из них мицелиев.
Другой также очень обычный плесеневый гриб, Aspergillus, принадлежит к высшим грибам и имеет членистый многоклеточный мицелий. На нем развиваются конидиеносцы в виде вертикально растущих ветвей длиной в 1— 2 мм, вздувающихся на конце. На поверхности этого вздутия образуются короткие цилиндрические выросты, отчленяющие на своих концах последовательно целые цепочки конидий (рис. 2). Последние отпадают и, попав на подходящий субстрат, легко прорастают, давая 2—3 ростковые трубочки, нарастающие концами и ветвящиеся. В отличие от Mucor, они вскоре делятся поперечными перегородками, так что получается многоклеточный мицелий. На нем опять вскоре формируются такие же конидиеносцы и т. д.
Рис. 2. Aspergillus. 1 — конидиеносец с цепочками конидий; 2 — половые органы; 3 — развитие плодового тела (спиральное тело, в середине — разросшийся женский половой орган с зачатками аскогенных гиф на нем); 4 — часть разреза через аскокарп с сумками и аскоспорами в них.
На том же мицелии развивается и половое спороношение. Вначале появляются половые органы в виде парами расположенных коротких ветвей, часто одной и той же гифы. Одна из этих ветвей, спирально закрученная, является женским органом, а другая, более прямая — мужским (рис. 2, 2). Вскоре затем женский орган дает боковые короткие выросты, называемые аскогеными гифами. На их концах формируются сумки, а в них сначала происходит слияние двух имеющихся ядер и затем редукционное деление копуляциояного ядра. После следующих затем еще двух делений в сумке оказывается восемь ядер, и около них в каждой сумке формируется по восемь аскоспор. Последние, как происходящие в результате редукционного деления, оказываются такими же неоднократными в генетическом отношении, как и споры Mucor в зародышевом спорангии.
Одновременно с указанным развитием половых органов происходит обрастание их соседними ветвями мицелия, которые плотно сплетаются в округлое тельце, и уже внутри него развиваются аскогенные гифы и сумки. Аскоспоры освобождаются вследствие разрушения оболочки этого тельца, называемого сумчатым плодовым телом, или аскокарпом (рис. 2, 4).
Третим и последним примером можно взять какой-нибудь из шляпных грибов, например часто развивающийся на навозе Coprinus (рис. 3). На его многоклетном мицелии могут развиваться, хотя и не особенно обильно, бесполые (конидиальные) спороношения, а затем залагаются плодовые тела, или базидиокарпы, состоящие из пенька и шляпки. На нижней стороне последней развиваются в очень большом числе спороносные органы — базидии. Так же как и сумки, они имеют сначала по два ядра, которые затем сливаются, и после этого копуляционное ядро делится редукционным путем. Затем следует еще одно деление, и в базидии получается четыре ядра. Тем временем на ее поверхности образуются четыре выроста, и в них переходит по одному ядру (рис. 3, 2). Таким образом, на каждой базидии образуется по четыре экзогенных споры, называемых базидиоспорами. По характеру своего развития, при котором происходит слияние клеточных ядер и затем редукционное деление, они сходны с аскоспорами и должны быть отнесены к половым спороношениям. Как те, так и другие, в отличие от бесполых спор, оказываются генетически неоднородными.
Риге. 3. Coprinus. 1 — плодовое тело с остатками диплоидного мицелия у основания ножки; 2 — базидии (b) с 4 базидиоспорами каждая.
Полиморфизм.
Из всех указанных органов — мицелия и органов спороношения — первый построен настолько однообразно, что идентификация гриба по нему большей частью совершенно невозможна. Характерные признаки приходится поэтому искать во вторых. Однако один и тот же гриб может иметь их несколько: половые спороношения и бесполые, притом последних нередко целый ряд (до 5—6) и иногда весьма различных. Благодаря этому гриб может в корне менять свою физиономию на разных стадиях, так что, если бы не знать генетической связи между ними, то их пришлось бы принять за совершенно разные организмы. Раньше обыкновенно так и делалось. Например, конидиальные спороношения разобранного выше в качестве примера Aspergillus назывались Aspergillus glaucus, а сумчатые — Eurotium herbariorum. Также для представителей современного рода ржавчинных грибов, Puccinia, одно спороношение носило название Spermogonium, другое — Aecidium, третье — Uredo, четвертое — собственно Puccinia, — и все это не в качестве стадий развития одного и того же организма, а в качестве самостоятельных даже родов. Позднее, при изучении полного цикла развития и установлении генетической связи спороношений, эти названия утратили прежний смысл и, если сохранились иногда, то лишь для обозначения того или иного спороношения (например, в приведенном выше примере ржавчинных грибов — названия Aecidium, Uredo и др.).
Это явление — способность грибов менять свой внешний вид в состоянии разных спороношений — получило название полиморфизма, или плеоморфизма их.
Значение для идентификации грибов разных спороношений.
Ввиду указанного полиморфизма встает вопрос, насколько то или иное спороношение может характеризовать данный гриб. Здесь приходится различать органы бесполого и полового спороношений. Поскольку дело касается первых (бесполых), то в некоторых случаях они вполне типичны для данного круга форм и не повторяются у других. Таковы, например, спорангии у мукоровых.
То же, пожалуй, хотя и не с такой точностью, можно сказать относительно конидиеносцев Aspergillus и некоторых других. Однако очень часто бесполые спороношения не достаточно характерны для установления систематического положения гриба. Таковы, например, конидиеносцы так называемого типа Oedocephalum, похожие отчасти на Aspergillus, но отличающиеся тем, что конидии сидят на вздутом кончике конидиеносца не цепочками, а в один только этаж. Они встречаются у Peziza vesiculosa из аскомицетов, у Fomes annosus из базидиомицетов (трутовиков), у Cunninghamella из мукоровых, у Basidiophora из пероноспоровых, а, кроме того, существует еще немало форм с такими же конидиеносцами, у которых неизвестны, кроме них, иные спороношения и которые причисляются поэтому к группе несовершенных грибов (Fungi imperfecti) (рис. 4, 2). Другой пример: похожие на Oedocephalum, но не вздутые на конце конидиеносцы типа Botrytis известны у многих несовершенных грибов (не имеющих, кроме них, других спороношений); кроме того, они развиваются как бесполое спороношение у сумчатого гриба Sclerotinia fuckeliana (рис. 4, 2), тогда как у многих других видов того же рода Sclerotinia бесполые спороношения совсем иного рода, в виде так называемой Моnilia (цепочка конидий на коротком конидиеносце). Если бы основываться здесь на бесполых спороношениях, то пришлось бы разбить очень характерный и хорошо отграниченый род Sclerotinia. Наоборот, половые спороношения оказываются очень характерными не только для идентификации данного гриба, но и для установления его филогенетических отношений. Поэтому они кладутся в основу изучения, и на основании их производятся систематические группировки.
Рис. 4. Конидиеносцы: 1 — типа Oedocephalum; 2 — типа Botrytis.
Половое воспроизведение.
Ввиду того, что половое воспроизведение грибов весьма своеобразно и у многих изменило свою первоначальную форму, для понимания необходимо вкратце остановиться здесь на его цитологической основе. Типичный половой процесс в основе своей имеет слияние двух клеток (гамет) и их клеточных ядер. Получившееся при этом копуляционное ядро содержит двойное число хромосом и называется диплоидным. В известный момент развития, разный для разных организмов, диплоидное ядро претерпевает редукционное деление, которым снова восстанавливается гаплоидное число хромосом. Таким образом, для каждого организма, имеющего половой процесс, характерно, что в его цикле развития имеется чередование гаплоидного и диплоидного состояний, или ядерных фаз (от копуляции до редукции — диплоидная фаза, от редукции до новой копуляции — гаплоидная).
Если подойти с этой точки зрения к половому воспроизведению грибов, то можно заметить следующее: у низших грибов (архимицетов и фикомицитов) непосредственным результатом оплодотворения является зигота, имеющая характер покоящейся споры. При прорастании ее копуляционное диплоидное ядро (или ядра, если как у Mucor оплодотворение заключалось в слиянии двух многоядерных клеток) делится редукционным способом, благодаря чему восстанавливается гаплоидное число хромозом. Таким образом, здесь диплоидная фаза представлена только покоящейся зиготой, и вся активная жизнь гриба проходит в гаплоидном состоянии.
У аскомицетов мы также наблюдаем оплодотворение. Однако при этом за слиянием половых клеток, или цитогамией (вернее плазмогамией) не следует сейчас же слияния их ядер, или кариогамии. Мужские и женские ядра в оплодотворенном женском половом органе (архикарпе) только сближаются попарно друг с другом, образуя так называемые синкарионы, или дикарионы.
При разрастании половых органов и развитии из них аскогенных гиф дикарионы размножаются, причем в ряде случаев прослежено, что оба ядра делятся строго одновременно и параллельно друг другу. В молодой сумке происходит слияние этих двух ядер дикариона в одно копуляционное диплоидное ядро. Оно затем сейчас же претерпевает редукционное деление, так что аскоспоры содержат уже гаплоидные ядра и прорастают в гаплоидный мицелий. Часть этого цикла развития от плазмогамии до кариогамии может быть отождествлена с диплоидной фазой, так как каждый содержащийся здесь дикарион ведет себя как нечто единое и содержит двойное число хромосом. Поэтому по существу является второстепенным моментом, что они заключены здесь в две ядерные оболочки, а не в одну. Таким образом, у аскомицетов чередование ядерных фаз выражается в следующем: от аскоспор до плазмогамии идет гаплоидная фаза; от плазмогамии до кариогамии, в сумке — диплоидная фаза, или, как ее называют еще по характерному строению ядерного аппарата, дикариофит. В отличие от низших грибов диплоидная фаза представлена здесь не покоящимся состоянием (зигоспорой), а имеет известное разрастание и размножение своих ядер. Однако преобладание в цикле развития имеет все-таки гаплоидная фаза, так как на ее долю приходится не только вегетативный мицелий, могущий неограниченно размножаться путем образования бесполых спороношений, но и значительная часть сумчатого плодового тела (вся оболочка его и часть внутренней массы, кроме аскогенных гиф и молодых сумок). Нужно отметить еще, что у многих аскомицетов слияния половых клеток совсем не происходит, но всегда тем или иным образом (апогамия) происходит образование дикарионов и развитие аскогенных гиф, так что характер смены ядерных фаз при этом по существу не изменяется.
Что касается, наконец, базидиомицетов, то у них совершенно неизвестно образования половых клеток. Базидиоспоры здесь, как и аскоспоры, гаплоидны. При прорастании они дают гаплоидный, или первичный мицелий, который может размножаться бесполым путем, но в общем имеет непродолжительное существование и вскоре заменяется диплоидным, или вторичным мицелием. Последний возникает в результате анастомозов и слияний гиф одного первичного мицелия или чаще двух разных. Через анастомоз переходит ядро из одной клетки в другую, в результате чего получаются первые двуядерные клетки с дикарионами. Они затем разрастаются в целый мицелий, сложенный из таких двуядерных клеток (при этом, конечно, дикарионы размножаются подобно тому, как и в аскогенных гифах). Этот вторичный мицелий, относящийся к диплоидной фазе (дикариофит), обычно имеет более продолжительное существование и часто бывает даже многолетним. За счет его полностью слагается все плодовое тело. В развива юпщхся на нем базидиях, так же как и в сумках, происходит, наконец, слияние ядер дикариона, и копуляционное ядро сейчас же делится затем редукционным образом, так что развивающиеся на базидии базидиоспоры содержат уже по одному гаплоидному ядру. Базидиоспора снова прорастает в первичный гаплоидный мицелий. Таким образом, и у базидиомицетов мы имеем ясную смену ядерных фаз: от базидиоспоры до возникновения первых дикарионов — гаплоидная фаза; от первых дикарионов через весь вторичный мицеЛий и все плодовое тело до базидий — диплоидная фаза, цитологически представленная, как и у аскомицетов, в виде дикариофита, но в отличие от них здесь преобладающая. Из приведенного видно, что базидиальное спороношение, подобно сумчатому стоящее в связи со сменой ядерных фаз (рис 5), является по существу выражением половой функции. Поэтому оно вместе с половыми спороношениями других грибов привлекает особое внимание миколога, так как здесь находят наиболее надежные указания на филогенетические отношения и эволюцию изучаемых групп.
Однако было бы односторонним ограничиваться только статическим изучением половых спороношений. Истинная природа всякого организма лучше выясняется в его динамике, особенно в индивидуальном развитии, или онтогенезе. Особенное значение имеет такой подход для грибов с их часто очень сложным циклом развития. Поэтому современное морфолого-систематическое изучение грибов, дающее в первую очередь основу для понимания их истинной природы и филогенетических взаимоотношений, ставит перед собой более сложные задачи: выяснение полного цикла развития с учетом цитологических явлений и сравнительное изучение не только строения, но и развития как вегетативных органов, так и различных спороношений, образующихся в той или другой фазе развития.
Настоящее, второе, издание «Микологии» сохраняет тот же план и те же основные установки, как и первое, опубликованное в 1933 г. Как и раньше, в нем ставится задача дать некоторый синтез знаний о грибах, не ограничиваясь их морфологией и систематикой, но обращая также значительное внимание на условия их жизни, значение в природе и хозяйстве и в связи с этим на те экспериментальные направления, которые приобретают все большее значение в современных микологических исследованиях.
Автор попрежнему полагает, что такое построение более соответствует запросам, предъявляемым у нас к микологии, притом не только со стороны ботаников, но и со стороны фитопатологов, агрономов, техников и др., чем те очень хорошие, но только морфолого-систематические сводки, какие появились за последние годы в Западной Европе и Америке (Gaumann, Gwynne-Vaug- han, Bessey). В противоположность им можно указать на старое классическое руководство Де-Бари (Vergleichende Morphologie und Biologie der Pilze, 1884), которое еще более 50 лет назад широко трактовало микологию: не только как морфологию и систематику грибов, но и как их биологию, понимаемую в широком смысле и с особенным ударением на явлениях паразитизма.
Посвящая настоящее издание памяти А. Де-Бари в связи с исполнившимся в этом году пятидесятилетием со дня смерти этого величайшего исследователя- миколога, автор хотел вместе с тем указать, что современное положение нашей науки опять приводит нас к той же широкой трактовке ее, как это было заложено в его классических сочинениях.
Понятно, что фактический материал, каким мы сейчас располагаем, значительно обширнее и не может быть поэтому изложен с той почти исчерпывающей полнотой, как это было сделано Де-Бари в его руководстве 1884 г. Наша книга и не претендует на исчерпывающее изложение. В основном — это руководство, стремящееся отразить только основные факты и главнейшие направления в современной микологии, но вместе с тем оно имеет в виду читателя, предъявляющего более глубокие запросы к этой дисциплине. Предполагается, что это будут главным образом студенты старших курсов, специализирующиеся в этой области, аспиранты и начинающие научные работники, притом не только ботаники и фитопатологи, но отчасти и агрономы, техники и др.
Для некоторых из этих категорий предполагаемых читателей было бы, вероятно, желательно иметь также побольше справочного и методического материала. Однако автор не решился расширить книгу в этом направлении из опасения слишком большого увеличения ее объема. Тем более, что мы имеем уже сейчас очень основательные справочники Н. А. Наумова по методике культуры и микроскопического изучения грибов. Также обильный справочный материал по некоторым вопросам можно найти в «Основах микологии» А. А. Ячевского (1933). Эта книга, остановившаяся из-за смерти автора на первом томе, касается только некоторых разделов общей микологии, но благодаря обилию справочного материала, содержит более 1000 страниц. Можно думать, что обработка в том же масштабе всей микологии потребовала бы, вероятно, по крайней мере, еще двух таких же томов. В наши намерения совершенно не входило составление такого огромного руководства.
При сохранении плана и общих установок первого издания тем не менее второе издание значительно переработано и расширено. Это коснулось не только исправления исключительно обильных опечаток первого издания, печатавшегося очень спешно и во многих листах почти без корректуры, но и исправлений и дополнений по существу в связи с данными новейшей литературы и необходимостью отразить некоторые новые течения, недостаточно выявленные и даже совсем отсутствовавшие в то время, когда изготовлялась рукопись первого издания (была окончена и сдана в издательство в 1931 г.). Также были приняты во внимание некоторые критические замечания по поводу первого издания, полученные от товарищей по науке, за что автор выражает им свою признательность. Впрочем, не все из их пожеланий могли быть выполнены. Так, указывалось на желательность расширения отделов по систематике пиреномицетов и несовершенных грибов. Этого не сделано, по крайней мере, в значительных размерах. Вполне соглашаясь, что каждый миколог должен знать значительное число представителей этих крупнейших групп, автор тем не менее полагает, что такие знания должны получиться из гербаризации и определения, чего не может заменить учебное руководство. Сделать же какие- либо существенные обобщения на основании более подробной классификации пиреномицетов и несовершенных грибов автор не видел возможности.
Как наиболее существенные изменения второго издания можно указать следующие:
В целом несколько смягчено догматическое изложение различных гипотез и толкований (особенно в вопросах филогенеза) приведением других существующих мнений на этот счет и их кратким критическим разбором. Расширена глава о гетероталлизме. Расширено изложение учения о биотипах и их возможном гибридном происхождении. Последнее сделано как в общей главе о паразитизме, так и более специально в главе о ржавчинных, где эти явления наилучше изучены. Переработана глава VIII — об общих условиях жизни грибов—и расширена изложением учения о значении ростовых веществ и биоса, а также явлений антагонизма. Расширено изложение учения о симбиозе. Расширены в систематической части главы о головневых и особенно ржавчинных грибах. Приводимые в конце глав списки литературы значительно расширены. Однако и в таком виде они отнюдь не могут претендовать на исчерпывающую полноту. Заново написан довольно подробный очерк истории микологии. Он приложен в конце книги, так как построен таким образом, чтобы показать развитие тех проблем, современное состояние которых излагается в основной части.
В общем, в результате указанных и многих других изменений и расширений общий объем текста увеличился примерно на 20—25%, а число иллюстраций — с 237 до 326.