Половое воспроизведение у грибов и связанные с ним процессы апогамии, смены ядерных фаз и пр. представляют много своеобразного и различного у разных групп. Ввиду того, что выяснение подробностей характера и эволюции полового процесса является важнейшим моментом дая понимания всей эволюции грибов, этим вопросам будет посвящено достаточное внимание в специальной части; здесь же можно ограничиться только освещением основных положений и схем.
Выше уже были отмечены основные различия в разбираемом отношении между низшими и высшими грибами. У первых (архимицетов и фикомицетов) непосредственным результатом оплодотворения является спора, имеющая почти всегда характер покоящейся споры. При прорастании она дает или прямо вегетативный мицелий (Monoblephaiis), или чаще развивает спорангий (так называемый зародышевый спорангий) со значительным количеством спор в нем (рис. 33, 5; 34, 4). У высших грибов (аскомицетов и базидиомицетов) продукт оплодотворения не переходит в состояние споры, а развивается дальше в особую диплоидную генерацию, которая в конце своего развития приносит обыкновенно в большом числе спорообразующие органы, а в них или на них — споры размножения. Результатом развития этой диплоидной генерации, цитологически характеризуемой как дикариофит (см. Половое воспроизведение), является увеличение числа спорообразующих органов и этим увеличение количества спор, которые имеют свои корни в половом акте и при образовании которых имеет место редукционное деление. А это генетически соответствует разъединению признаков и образованию новых комбинаций их, т. е. получившиеся таким путем споры оказываются генетически неоднородными.
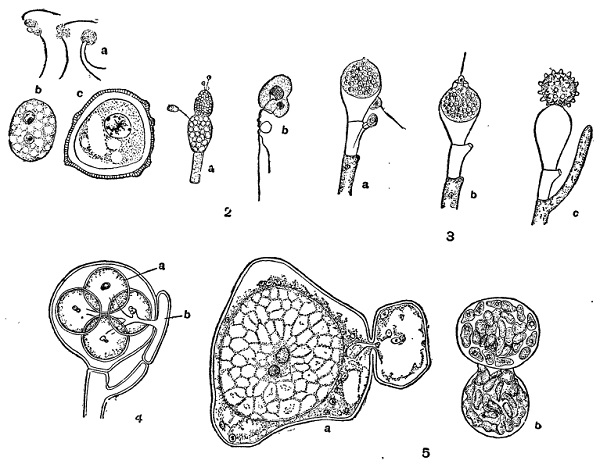
Рис. 33. Половой процесс низших грибов (архимицетов и оомицетов). 1 — Olpidium viciae: a — копуляции гамет; Ь и с — молодая и взрослая зиготы. 2 — Allomyces javanicus: а — мужской и женский гаметангии; b — копуляция гамет. 3 — Monoblepharis sphaerica: а и b — оплодотворение; с — ооспора. 4 — Saprolegnia: а — яйцеклетки; b — антеридий. б — Albugo Candida: a — оплодотворение; b — прорастание ооспоры.Рис. 34. Половой процесс зигомицетов. 1 — Rhizopus nigricans; 2 — Absidia spinosa — гетерогамная копуляция; 3 — Endogone — гетерогамная копуляция одноядерных клеток; 4 — Мuсоr—прорастание зигоспоры образованием зародышевого спорангия.
Бесполое размножение свойственно преимущественно гаплоидной фазе грибов, поэтому оно очень типично для фикомицетов, у которых вся жизнь за исключением зиготы проходит в гаплоидном состоянии, и широко распространено у аскомицетов, где также гаплоидное состояние является преобладающим.
Рис. 32. 1 — эндоконидии у Thielavia; 2 — Nyctalis, хламидоспоры между базидиями.
Сюда же нужно причислить и несовершенные грибы как большею частью гаплоидные бесполые состояния сумчатых. Что касается базидиомицетов, у большинства которых преобладает диплоидная фаза, а гаплоидная представлена только большею частью недолговечным первичным мицелием, то в соответствии с этим бесполое размножение у них за исключением Uredinales распространено значительно меньше и не достигает такого разнообразия и совершенства форм. Наиболее сложные бесполые спороношения — пикниды — здесь почти совсем неизвестны (за исключением относящихся к гаплоидной фазе спермогониев Uredinales). Главным образом здесь наблюдаются несложно построенные конидиеносцы без образования цепочек конидий или еще чаще оидии, относящиеся уже к вегетативному размножению. Один из наиболее совершенных примеров органов бесполого размножения базидиомицетов мы имеем у Fomes annosus, где они представлены конидиеносцами типа Oedocephalum, шаровидно вздутыми на конце и несущими многочисленные конидии. Органы бесполого размножения у высших базидиомицетов образуются, насколько известно, преимущественно на первичном гаплоидном мицелии, на диплоидном же известны главным образом хламидоспоры (например у Nyctalis из Agaricaceae, а также у некоторых Polyporaсеае, рис. 23, 2). Диплоидны также и упомянутые конидиеносцы Fomes annosus.
Рис. 23. Гаустории пероноспоровых. 1 — Albugo; 2 — Peronospora parasitica; 3 — то же в поперечном разрезе; видно положение разрезанной гифы в межклетнике и отхождение от нее гаусторий в две клетки.
Выше было разобрано строение отдельных кониеносцев; однако у многих форм они соединяются в тесные группы, которые делят обычно на следующие три основные категории: лоремия, ложе и пикнида. Впрочем, резкой границы между ним, провести нельзя.
Коремия
Коремией называют пучок тесно сближенных и склеенных большей частью своими боками конидиеносцев, отшнуровывающих каждый на своей верхушке конидии. Они характерны особенно для целой группы несовершенных грибов, называемой Coremiales (представители, например, Stysanus, Graphium, Sphaerostilbe и др., рис. 30, 2), но встречаются изолированно и среди других групп, например у некоторых видов Penicillium. Типичные коремии имеют вид узких и сравнительно высоких колонок; если они более широкие и низкие, то представляют переход к ложу.
В типичном случае состоит из плоского сплетения гиф, на поверхности которого тесным слоем, называемым иногда гимением, располагаются конидиеносцы, большей частью короткие и мало или совсем не ветвящиеся. На концах своих, т. е. на верхней свободной поверхности своего слоя, они отделяют конидии (большей частью одиночные). Такие ложа особенно характерны для большой группы несовершенных грибов Меlanconiales (представители: Gloeosporium, Goryneum, рис. 30, 2), но встречаются и в других группах, например в уредо-спороношениях ржавчинных грибов. Часто ложа залагаются внутри субстрата (ткани питающего растения у паразитных форм) и долгое время остаются прикрытыми, освобождаясь лишь через разрыв наружных покровов к моменту созревания спор. Нередко, далее, с боков ложа окружены кольцом стерильных гиф, представляющих зачаточную оболочку всего спороношения. При большем развитии ее получается переход к пикниде.
Пикнида
В типичном виде представляет округлое образование, одетое общей параплектенхиматической или прозоплектенхиматической оболочкой с узким отверстием наверху. От оболочки внутрь по радиусам отходят расположенные тесным слоем короткие конидиеносцы. Отделяемые ими споры (пикноспоры или стилоспоры) скопляются в центральной полости и оттуда через верхнее отверстие выходят наружу, погруженные обыкновенно в общую массу слизи. Пикниды типичны для большой группы несовершенных грибов Spaeropsidales (Septoria, Phoma и др., рис. 30, 3) и встречаются также в качестве конидиальных спороношений у многих сумчатых грибов, а также у ржавчинных (так называемые спермогонии). В подробностях строение пикнид может быть различным. Иногда они не одиночны, а соединены по нескольку в общей строме; иногда не округлены, а приплюснуты сверху вниз и при созревании открываются более широким отверстием и т. д. Последнее является переходом к ложу.
Эндоконидии
От типичных экзогенных конидий несколько отличаются так называемые эндоконидии. Они известны у немногих форм (Thielavia, Endoconidiophora и некоторых других) и образуются следующим образом: прямые неветвящиеся конидиеносцы, начиная сверху, делятся на короткие клетки-конидии. По мере их развития и обособления внутреннего, слоя оболочки от наружной общей боковой стенки конидиеносца они постепенно выталкиваются из нее, как из чехла, растущими нижними клетками. Таким образом, в основе мы имеем здесь ту же цепочку конидий, но только в ее образовании не принимают участие самые наружные слои оболочки (рис. 31 и 32). Подобные же по своему происхождению эндоконидии развиваются у Laboulbeniales, но там им приписывается функция мужских половых элементов, почему их называют спермациями.
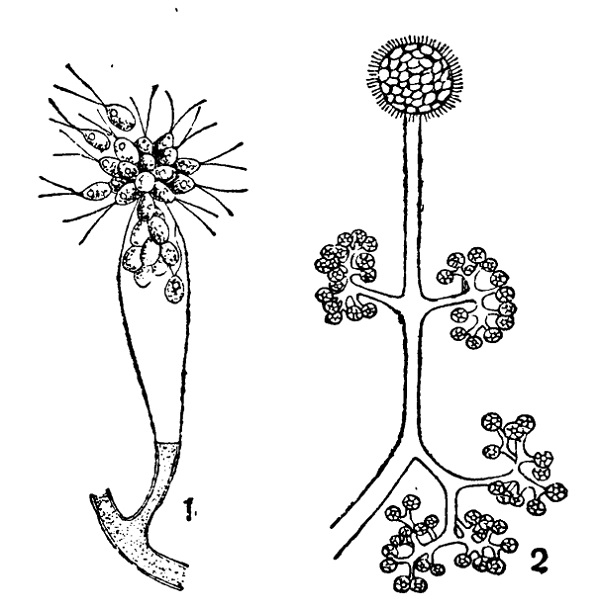
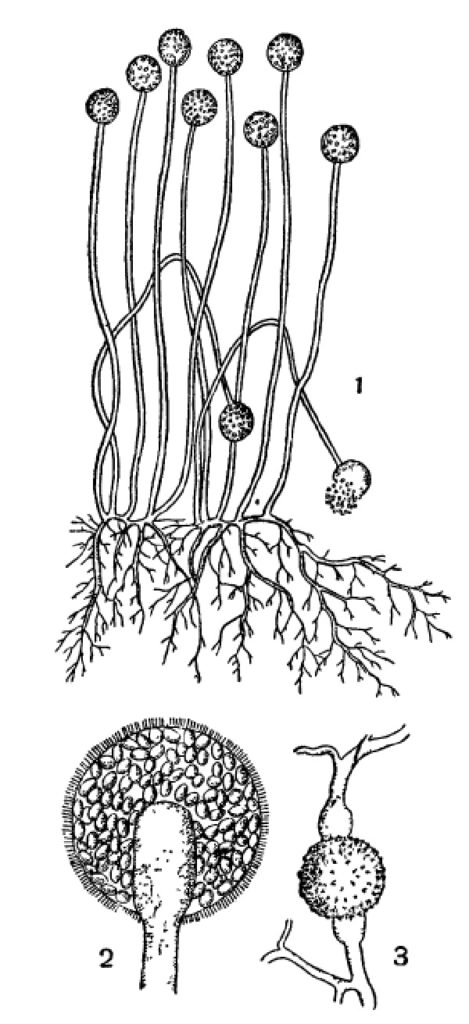
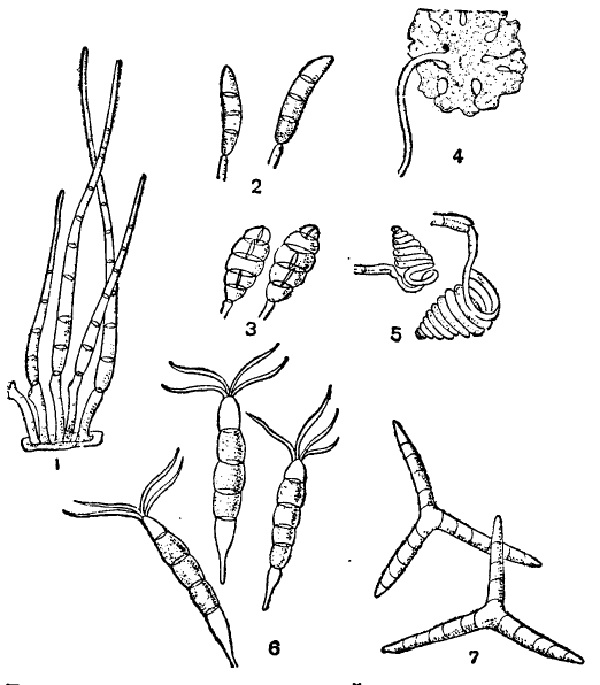
Что касается развития конидий, то оно происходит следующим образом. Кончик конидиеносца слегка разрастается и дает обыкновенно небольшое вздутие, куда притекает особо обильная протоплазма с клеточными ядрами. Затем появляется поперечная перегородка, отделяющая вздутый кончик от нижележащего конидиеносца. Конидия затем увеличивается в размерах, утолщает свою оболочку отложением внутренних слоев, первичная же поперечная перегородка очевидно расслаивается затем на две пластинки, которые отклеиваются друг от друга, начиная от периферии, и выгибаются от тургора обеих клеток наружу, навстречу друг другу. Лишь в самом центре, где находится протоплазменное соединение между конидией и конидиеносцем, связь остается дольше. Когда обрывается и она, конидия спадает. В некоторых случаях этим образованием одиночной конидии дело и кончается, но чаще после опадения первой конидии подобным же образом на ее месте образуется вторая и т. п. Это имеет место, например, в большинстве пикнид, а также и среди многих свободных конидиеносцев (Verticillium и многие другие, рис. 29,2). Не менее часто наблюдается, что на конце конидиеносца одновременно находится несколько конидий, или расположенных головкой или в ряд — четкообразно. Первое — образование головки — происходит таким образом, что на конце конидиеносца последовательно или одновременно образуется несколько выростов, иногда ничтожной величины, которые, однако, каждый отчленяют вышеописанным образом по конидии. При этом нередко кончик конидиеносца головчато расширяется, чем увеличивается поверхность для образования большего числа конидий (Oedocephalum, рис. 29, 2). У Aspergillus выросты, отчленяющие конидии, имеют характер небольших многоядерных цилиндрических клеток, соединенных в основании узким отверстием с общей полостью головки конидиеносца; они называются стеригмами. У близкого к Aspergillus рода Sterigmatocystis дело пошло еще дальше: здесь каждый такой вырост состоит из основной клетки, а на дистальном конце ее располагается несколько цилиндрических стеригм, несущих конидии (рис. 29, 3). Образование головки, особенно в таких случаях, как у Aspergillus или Sterigmatooystis, может рассматриваться, строго говоря, как образование коротких ветвей на конце конидиеносца (аналогично, например, некоторым соцветиям, как головки или корзинки сложноцветных); однако при описании соответственных форм их конидиеносцы определяются обычно как неразветвленные, так как эдесь принимаются во внимание только более развитые ветви.
Четка, или цепочка, конидий может быть базипетальной или акропетальной. Первая развивается следующим образом: после отделения первой конидии, но раньше чем она отпадет, конидиеносец под ней подобным же образом развивает вторую, за ней третью и т. д. В результате получается цепочка, в которой верхняя конидия — самая старая и обыкновенно самая крупная, а от нее вниз идут все более молодые и мелкие. По мере того как верхние конидии обваливаются, снизу, со стороны стеригмы, образуются новые, так что цепочка в общем сохраняет свою длину. Этот тип наиболее обычен. Он прекрасно выражен, например, у Erysiphaceae (рис. 29, 5) или на более сложном разветвленном конидиеносце у Penicillium (рис. 29, 4) и многих других. Акропетальные цепочки встречаются значительно реже. При их развитии первая отделенная конидия, разросшись и сделавшись большей частью многоклетной, дает на своей вершине вырост, развивающийся дальше в такую же вторую конидию. На ее вершине таким же образом развивается третья и т. д. (рис. 29, 6). В результате в цепочке самой старой и крупной оказывается нижняя конидия, а самой молодой — верхняя. Акропетальные цепочки могут иногда ветвиться, что происходит благодаря появлению на верхушке какой-нибудь конидии не одного, а двух выростов, развивающихся указанным образом в две цепочки. В базипетальных цепочках, как ясно из их развития, таких ветвлений быть не может. Акропетальный тип встречается большей частью у форм с сложными многоклетными конидиями (Septosporium, Alternaria).
У Aspergillus, Sterigmatocystis и некоторых других каждая стеригма, сидящая на головке, отделяет не одну конидию, а целую базипетальную цепочку. Таким образом, здесь скомбинированы и головчатое и четкообразное расположение спор, и головка является сложной (подобно, например, какому-нибудь сложному соцветию). Конечно, благодаря этому она несет особо большое число конидий. Например, по приблизительному подсчету у Sterigmatocystis niger в одной головке содержится их не менее 10 000; кроме того, нужно принять во внимание их возобновление по мере отпадения более старых. Из других сложных и особенно типичных конидиеносцев следует указать еще Penicillium. Здесь многоклетный конидиеносец в верхней части дает две-три ветви, которые в свою очередь два-три раза ветвятся, причем последние разветвления представляют стеригмы, а на них сидят базипетальные четки спор. Все спороношение имеет известное сходство с кисточкой, откуда и название Penicillium (рис. 29, 4).
Как было указано выше, под бесполым размножением мы понимаем такое, которое осуществляется при посредстве особых спор. Последние развиваются обыкновенно не нецосредственно на мицелии, а на особых более или менее резко обособленных ветвях его. В свою очередь эти споры могут быть эндогенного и экзогенного происхождения. Первые образуются обыкновенно в большом числе внутри особых клеток — спорангиев, почему самые споры называются спорангиоспорами, вторые (экзогенные) образуются на поверхности производящих их органов, которые называют конидиеносцами, а самые споры — конидиями. У низших грибов преобладает образование спорангиоспор, и только у сравнительно немногих встречаются конидии, тогда как у высших имеются только последние. Причину этого явления нужно видеть в общей эволюции грибов, шедшей в сторону приспособления к наземному образу жизни. Из низших форм многие являются еще чисто водными растениями, и даже их споры нередко представлены в виде зооспор. Однако из них уже у мукоровых грибов, перешедших к наземному существованию, наблюдается местами превращение спорангиев в конидии. Можно думать, что в воздушной среде не всегда имеются достаточно благоприятные условия (например влажность) в течение того сравнительно значительного промежутка времени, какой требуется для развития массивного спорангия; маленькие же конидии могут быстро закончить свое развитие и этим обеспечить размножение гриба. У высших грибов, еще более приспособленных к наземному существованию, конидии сделались единственным элементом бесполого размножения.
Эндогенные спороношения.
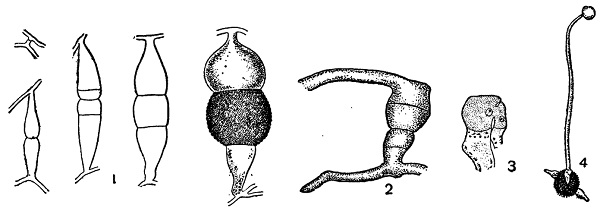
Как было указано выше, у некоторых низших грибов в спорангиях образуются зооспоры. Они представляют голые участки протоплазмы с одним ядром и одним или двумя жгутиками, при помощи которых активно передвигаются в воде. Такие зооспоры характерны для архимицетов и оомицетов (рис. 27, 1), Спорангии, в которых они образуются (зооспорангии), большею частью диференцированы весьма слабо и развиваются из кончиков обыкновенных вегетативных гиф. Они отделяются перегородкой от остальной части гифы, и зооспоры образуются в них в результате распадения общей протоплазмы на голые одноядерные участки, вырабатывающие жгутики. У других грибов (главным образом Мucoraceae) споры представляют клетки, одетые твердой оболочкой, лишенные органов движения и большей частью многоядерные. Спороношения здесь более высокодиференцированы. Спорангии образуются на особых спорангиеносцах, которые, хотя и являются по происхождению ветвями вегетативного мицелия, но резко отличаются обыкновенно своей большей толщиной, ограниченным ростом, характером ветвления (или полным отсутствием ветвления) и своими тропизмами (отрицательный геотропизм, положительный фототропизм и т. д.), благодаря чему растут кверху от субстрата (рис. 27, 2).
На их концах только в воздушной среде образуются спорангии в виде довольно массивных шаровидных вздутий, отделяющихся от спорангиеносца перегородкой. В yих уже формируются споры путем распадения общей протоплазмы спорангия на отдельные большей частью многоядерные участки, которые округляются и одеваются собственными оболочками (рис. 1, 1 и 2). Освобождаясь через разрыв оболочки спорангия, они распространяются далее токами воздуха.
Рис. 1. Mucor mucedo. 1 — спорангиеносцы со спорангиями и мицелий; 2 — спорангий, сильно увеличенный; 3 — зигоспора на месте слияния двух одинаковых ветвей мицелия.
Экзогенные (конидиальные) спороношения
Эти спороношения гораздо разнообразнее эндогенных. Во-первых, здесь различны самые споры или конидии. Они залагаются всегда как одна клетка, насколько известно, чаще многоядерная и имеющая большею частью округлую или овальную форму. Часто они и в дальнейшем остаются такими же (например у Penicillium, Aspergillus, Botrytis и многих других), но нередко они затем значительно вытягиваются в длину, принимая цилиндрическую или веретеновидную форму, делясь при этом обыкновенно поперечными перегородками на ряд клеток (у Cercospora, Fusarium, Septorian др.; рис. 28, 1 и 2). В других случаях дело не ограничивается только этими перегородками, а появляются и продольные, так что готовая конидия представляет уже довольно массивное клеточное тело, как у Macrosporium (рис. 28, 3). Встречаются и более сложные формы конидий, например в виде лопастных тел, напоминающих отчасти очертания клетки десмидиевых водорослей (Desmidospora), в виде нити, закрученной в крутую спираль (Helicosporium или Helicoon), в виде якоревидных тел (Trinacrium) и т. п. (рис. 28, 4, 5, 7). В некоторых случаях на одном или обоих концах веретеновидных конидий помещаются пучки длинных нитевидных выростов (Pestalozzia, Dilophospora) и т. д. (рис. 28, 6). Вообще наиболее сложно построенные конидии встречаются у так называемых Fungi imperfecti, где они являются единственными органами размножения.
Конидиеносцы, так же как и спорангиеносцы, представляют особые ветви мицелия, отличающиеся строением, характером ветвления и роста. Они поднимаются от субстрата кверху и образуют на своих концах конидии,за немногими исключениями, только в воздушной среде. Редко конидии образуются сбоку на конидиеносце, например у Tomentella.
Конидиеносец иногда состоит из одной клетки и тогда обыкновенно не ветвится (Aspergillus), но иногда и в этом случае наблюдается ветвление (Oedocephalum, рис. 29, 1). Более распространены многоклетные и притом разветвленные конидиеносцы. Их ветвление весьма разнообразно и характерно для отдельных групп и их представителей. Благодаря ему увеличивается продукция конидий, образующихся как правило только на концах ветвей (апикально).
Отделенный кусок грибницы как многоклеточной, так даже и неклеточной, обыкновенно легко регенерирует, помещенный в подходящие условия, и продолжает дальше свое развитие. Этот наиболее простой способ вегетативного размножения грибов несомненно очень распространен у них в естественной обстановке и практикуется также при искусственном разведении, например при культуре шампиньона. Несколько более специализированы такие также очень распространенные случаи, когда сама грибница благодаря особенностям своего строения легко разъединяется на отдельные клетки, которые дальше вырастают каждая в новую такую же грибницу. Сюда относится почкование, так как почкующийся мицелий параллельно со своим ростом распадается на отдельные части, т. е. размножается. Аналогичное вегетативное размножение у типичного гифенного мицелия осуществляется всего чаще путем образования оидий. Они получаются таким образом, что в гифах, начиная от концов их, появляются более частые чем обычно перегородки, и по ним происходит распадение на отдельные клетки большею частью овальной формы (откуда и название оидии, от греческ. оод — яйцо). Такое распадение может в конце концов захватить весь мицелий или ограничивается только его конечными разветвлениями. Отделившиеся оидии при подходящих условиях прорастают в новый мицелий. Образование их встречается у различных грибов, например у Endomyces и некоторых других Protascales, у многих гименомицетов и пр. (рис. 26, 3). Другими органами вегетативного размножения мицелия являются хламидоспоры. Они образуются, например, очень легко у Mucor racemosus при неблагоприятных условиях роста. При этом содержимое гиф стягивается в некоторые места и здесь одевается собственной толстой оболочкой. Это и есть хламидоспоры (рис. 26,4). После разрушения оставшихся пустыми частей гиф они освобождаются и могут прорастать в новый мицелий. Вместе с тем они могут выдерживать высыхание и действие других неблагоприятных условий, так что служат также для сохранения вида в течение неблагоприятного периода. В многоклетных мицелиях в хламидоспоры превращаются или целые вегетативные клетки, или предварительно разделившиеся на более короткие отрезки. В них скопляются запасные вещества, они округляются и одеваются под прежней оболочкой новой, более толстой и окрашенной. Хламидоспоры распространены у очень многих грибов, например у Perisporiaceae и многих других аскомицетов. Также у Ustilaginales их головневые споры по образованию являются хламидоспорами, хотя по характеру прорастания имеют особое значение (см. в спец. части Ustilaginales).
В случае, если клетки характера хламидоспор отличаются неопределенностью и разнообразием своей формы, то их называют геммами. Они встречаются как у низших грибов с неклеточным мицелием (Saprolegniales, Peronosporales), так и у высших.
Резкой границы между этими двумя типами размножения у грибов провести невозможно, однако все же такое разделение представляет известные удобства для обзора.
Под именем вегетативного размножения понимается такое, которое осуществляется не специализированными или мало специализированными частями вегетативного тела, т. е. грибницы. Этим оно противопоставляется бесполому размножению, осуществляемому при посредстве специальных спор. В свою очередь и то и другое очень резко противополагается половому размножению, которое также сопровождается образованием спор, но в основе своей имеет половой процесс, отсутствующий как при вегетативном, так и при бесполом размножении.
За исключением Myxochytridiales вегетативное тело у всех остальных грибов представлено мицелием, сложенным из системы ветвящихся гиф, растущих своими концами. У большинства низших грибов гифы не имеют поперечных перегородок, и весь мицелий является, формально говоря, одной сильно разветвленной многоядерной клеткой, достигающей нередко значительных размеров (10 см и больше между кончиками противоположных ветвей). Такой мицелий называется нечленистым, или одноклетным, или неклеточным. У всех высших грибов и некоторых низших в гифах мицелия на более или менее одинаковых расстояниях расположены поперечные перегородки, делящие их на отдельные клетки — одноядерные или многоядерные. Такой мицелий называется членистым, или много к летным.
От типичного гифенного мицелия отличается почкующийся мицелий. Он характеризуется тем, что его клетки размножаются не обычным делением, а почкованием и не остаются при этом прочно соединенными друг с другом. При почковании на поверхности клеток появляется выпуклина, или почка, которая постепенно увеличивается в размерах и отграничивается затем поперечной перегородкой от производящей клетки. На ней в свою очередь развиваются такие же почки и т. д. (рис. 26, 1). В результате получается непрочное соединение большей частью круглых или овальных клеток, которое и представляет почкующийся мицелий. В некоторых случаях он остается очень коротким (из 2—3 клеток), так как все время распадается на отдельные клетки, но в других — достигает больших размеров и ветвится. Почкование встречается у весьма многих и различных грибов. Наиболее типично оно для дрожжей и близких к ним форм, где происходит при всех условиях роста, для многих же других оно наблюдается только при некоторых специальных условиях, например при недостатке кислорода, избытке или, наоборот, недостатке питательных веществ. Почкование можно наблюдать у Mucor racemosus (рис. 26, 2), многих аскомицетов (Taphrina, Bulgaria, многих Sphaeriaeeae), большинства головневых и Tremellales, некоторых холобазидиомицетов (Exobasidmm) и ряда несовершенных грибов.
Из особых видоизменений мицелия нужно отметить: тяжи, ризоморфы, пленки, или мицелиальную кожу, и склероции.
Мицелиальные тяжи
Очень часто образуются у представителей самых различных групп высших грибов. В простейшем случае они сложены из небольшого количества параллельно идущих гиф, которые или просто склеиваются друг с другом ослизненными наружными оболочками, или вступают в более прочное соединение через посредство многочисленных коротких анастомозов (рис. 24, 1). Такие тяжи, различимые только в микроскоп, сложены из однородных гиф. В других случаях, когда они более массивны, их гифы получают известную диференцировку, выражающуюся чаще всего в том, что наружные элементы, более тонкие, образуют как бы кору, а внутренние, более широкие, — сердцевину, или центральный цилиндр. Наружные части коры часто особенно прочны и окрашены в тёмный цвет, тогда как середина остается светлой. Впрочем, бывают и обратные отношения: например в тяжах Phallus impudicus кора снежно-белая, а средняя часть бурая. Наибольшей диференцировки достигает строение тяжей у домового гриба (Merulius lacrymans).
Рис. 24. 1 — мицелиальный тяж гриба «черни»; 2 — ризоморфа опенка с плодовыми телами на ней: а и Ь — плодовые тела; г — ризоморфа.
Здесь в них можно различить:
обыкновенные тонкостенные гифы;
гифы с узким просветом и сильно утолщенными оболочками (механические);
гифы с очень широким просветом, неравномерным внутренним утолщением боковых стенок и разрушенными отчасти поперечными перегородками (сосудистые, или проводящие гифы).(рис. 6, 2).
Рис. 6. 1 — Merulius, гифы из тяжа, Н — обыкновенные вегетативные гифы; F — механические гифы с утолщенными стенками, G — сосудистые гифы с неравномерно утолщенными стенками; 2 — Battarea, клетка капиллиция; 3 — Mutinus caninus, гифа с сферокристаллом оксалата внутри и отдельными кристаллами на поверхности.
Ризоморфы
Более массивные тяжи с темноокрашенными наружными» частями имеют известное сходство с корнями высших растений и получили название ризоморфы. Особенно типично они представлены у опенка (Armillaria mellea), где собственно и дано было сначало это название — ризоморфа. Тяжи и ризоморфы нарастают, как и слагающие их гифы, своими вершинами и могут достигать нескольких миллиметров толщины и нескольких метров длины (рис. 24, 2). Они распространены у очень многих грибов с крупными плодовыми телами как у почвенных сапрофитов, так и у древесных форм: из базидиальных — у Phallaceae, Lycoperdaceae, Nidulariaceae, Agaricaceae, Polyporaceae; из сумчатых — у Elaphomyces, Genea, некоторых Peziza и др.
Роль тяжей и ризоморф состоит отчасти в том, что благодаря своему значительному разрастанию они способствуют вегетативному распространению гриба (например у опенка — от дерева к дереву часто на несколько метров); с другой стороны, через них может происходить передвижение веществ. Например, у домового гриба передвижение особенно обеспечивается наличием сосудистых гиф, благодаря чему гриб может перекидываться на значительные расстояния. Также и тяжи у основания крупных плодовых тел Phallus и др. служат для подведения к ним питательных веществ из мицелия. Наконец, ризоморфы, защищенные плотной корой, могут служить и для сохранения гриба при неблагоприятных условиях.
Мицелиальная пленка, или кожа (Myzelhaut)
Особенно типична для различных грибов, разрушающих древесину. Они нередко образуют в трещинах субстрата или на его свободной поверхности слои плотно переплетенных гиф до нескольких миллиметров толщиной, имеющих внешнее сходство с замшей. В дальнейшем на них развиваются плодовые тела или от них отходят тяжи, или, наконец, простой мицелий.
Рис. 25. 1 — склероции Polyporus sapurema с плодовым телом на нем (уменьш.); 2 — склероции Sclerotinia urimla с плодовыми телами.
Склероции
Под этим именем известны более или менее темные округлые тела плотной консистенции, образованные в результате тесного переплетения и анастомозов гиф. По размерам склероции бывают от десятых долей миллиметра до 30 см в диаметре и 20 кг веса. Наиболее крупные склероции имеют некоторые виды тропических трутовиков, например Polyporus sapurema из Бразилии — до 30 см в диаметре (рис. 25, 2), Pol. mylittae из Австралии — величиной с голову человека, Pol. sacer из юго-восточной Азии — величиной с кулак. Анатомическое строение склероциев довольно однообразно: снаружи лежит кора из одного или чаще нескольких слоев клеток с темными стенками, плотно соединенными друг с другом; нутри находится белая сердцевина, имеющая характер или параплектенхимы, или прозоплектенхимы. И то и другое может встречаться даже в одном склероции, например у спорыньи наружные части построены по первому типу, а внутренние — по второму. В зрелом состоянии склероции очень бедны водой (5—10%) и содержат значительное количество запасных питательных веществ, особенно жира (например в спорынье до 30% жира). В таком виде они могут сохраняться долгое время и затем при благоприятных условиях прорастают или прямо в плодовое тело, или развивая мицелий. Склероции распространены среди всех групп высших грибов: из аскомицетов, например, у Claviceps purpurea (спорынья) и особенно у рода Sclerotinia, которая даже получила свое название от этого образования (рис. 25, 2); среди базидиальных грибов — у Typhula, некоторых видов Goprinus, Lepiota, Polyporus и многих других. Также встречаются они у представителей несовершенных грибов (Fungi imperfecti). Они образуются чаще всего свободно на мицелии, а у некоторых паразитных форм — внутри пораженного растения или в каких-нибудь полостях его, например у Sclerotinia libertiana часто в сердцевинной полости стеблей различных травянистых растений, или, наконец, мумифицируют целые органы, как, например, Sclerotinia vaccirui — ягоды черники и др. К тому же типу склероциев по анатомическому строению и физиологическому значению относятся развивающиеся у многих паразитных сумчатых грибов плоские подстилки под конидиальными спороношениями (так называемые ложа, или стромы; см. стр. 34). После зимовки они развивают сумчатые плодовые тела.