Обнаруживает больше разнообразия. Ветвление гиф обычно обильное, но иногда оно выражено слабо. Например, этим отличаются толстые гифы сапролегниевых и Monoblepharidaceae. Иногда наблюдается известная диференцировка гиф: одни обычно более толстые и слабо разветвленные, а другие — сильно ветвящиеся и более тонкие. Первые служат, главным образом, для распространения мицелия по субстрату, а вторые для воспринятия из него в подходящих местах питательных материалов. Такая диференцировка особенно характерна для мицелия некоторых паразитных грибов, но встречается нередко и среди сапрофитных: например у Rhizopus и некоторых других имеются особые столоны, не ветвящиеся и обладающие энергичным ростом. Они совершенно аналогичны таким же столонами («усам»), например, у земляники и служат также для быстрого вегетативного распространения по субстрату. У таких снабженных столонами форм имеются еще нередко особые органы прикрепления — так называемые ризоиды — в виде пучка коротких разветвленных нитей. Они развиваются как реакция на соприкосновение с твердым субстратом, притом не только с таким, который может иметь известное питательное значение, но и например со стеклом (рис. 22, 1). У некоторых видов Sclerotinia названные органы прикрепления имеют вид довольно массивных кисточек из пучка тесно переплетенных коротких ветвей (рис. 22, 2); то же на неклеточном мицелии наблюдается, например, у Gircinella minor.
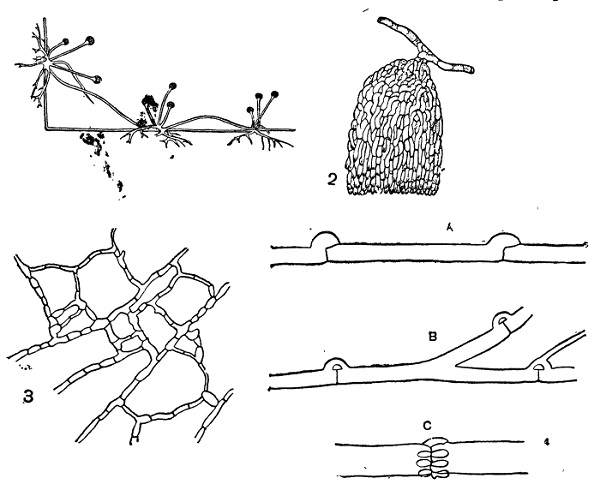
2 — Sclerotinia tuberosa, органы прикрепления мицелия;
3 — анастомозы мицелия;
4 — пряжки: А — развитие пряжки; В — пряжки Merulius; С — Coniophora.
При обильном ветвлении гифы мицелия соприкасаются друг с другом и здесь, на месте соприкосновения, нередко происходит слияние их с установлением сообщения между клетками. Такие слияния называются анастомозами; при большом развитии их мицелий приобретает характерный сетчатый вид (рис. 21 и 22, 3). Развитие анастомозов наблюдается у весьма различных грибов с многоклетным мицелием. В некоторых случаях через них совершается перемещение клеточного ядра из одной клетки в другую, и осуществляется переход от гаплоидного к диплоидному мицелию (у многих гименомицетов,см. стр. 56). Однако большей частью они имеют чисто вегетативное значение и развиваются у многих форм просто при недостатке питания.

1 — анастомозы, черные — две проросшие базидиоспоры;
2 — начало образования мицелиального тяжа;
3 — клетка, мицелия с окрашенными клеточными ядрами.
В некоторых случаях гифы снабжены еще так называемыми пряжками, представляющими небольшие плоские клетки, лежащие сбоку против поперечной перегородки и сообщающиеся в готовом состоянии открытым отверстием с полостью нижней (проксимальной) клетки. Такие пряжки встречаются только у базидиальных грибов, главным образом у гименомицетов, хотя и не у всех. Их наличие может служить важным признаком для установления принадлежности данного мицелия именно к этой группе. В деталях в свою очередь пряжки бывают различны. Они могут быть или против каждой поперечной перегородки или только против некоторых. Большею частью здесь образуется только одна пряжка, но, например, у очень распространенного разрушителя древесины в постройках, Coniophora cerebella, они развиваются целой мутовкой в количестве до 10 и более штук (этот признак весьма существенен при диагносцирова
нии ConiopKora по стерильному мицелию, в каком виде она всего чаще встречается). Наконец, в ряде случаев наблюдается так называемое прорастание пряжек, т. е. развитие из них боковой ветви (например у Merulius и некоторых других, рис. 22, 4b). Пряжки развиваются исключительно на диплоидном (двуядерном) мицелии и служат аппаратом, через посредство которого происходит передача ядра из одной клетки в другую (см. Basidiomycetes, стр. 310). Кроме многих гименомицетов, они встречаются в диплоидном мицелии иногда Ustilaginales и, по некоторым указаниям, также у Uredinales.