Стебель виноградного растения — это осевой вегетативный орган, соединяющий листья с корневой системой. Он выполняет рад важнейших функций: передвижение питательных веществ и воды от корней к листьям, транспорт продуктов ассимиляции вниз, накопление запасных питательных веществ, удаление и накопление вредных веществ, размещение листового аппарата, опорную функцию. В тканях стебля осуществляется ряд жизненно важных процессов метаболизма растения. В процессе развития стебля происходит освоение виноградным растением надземного жизненного пространства. Части стебля вместе с почками участвуют в выполнении функции вегетативного (бесполого) размножения.
Стебель является осевой частью побега винограда. Побег представляет собой сочетание стебля с листьями, почками, соцветиями (гроздями), пасынками и усиками, расположенными на нём.
В естественном состоянии стебель винограда имеет большую длину (до 30 м) и небольшую толщину, он гибкий, быстро растёт. Строение стебля асимметричной формы. Все органы побега развиваются на узлах.
При выращивании в промышленной культуре виноградному растению чаще придают вид приземистого куста высотой около 2 м. Скелетная часть стебля куста включает в себя следующие органы: надземный и подземный штамбы, голову, многолетние рукава, ответвления рукавов, рожки, однолетние вызревшие побеги, обрезанные на определённую длину (плодовые побеги и сучки). Штамб (подземный и надземный), голова и рукава составляют многолетнюю древесину куста винограда. Однолетняя древесина (плодовые побеги и сучки) размещается, как правило, на двулетней древесине рожков.
На скелетной части куста ежегодно весной из перезимовавших почек развиваются основные побеги текущего года: плодоносные и бесплодные — из почек зимующих глазков на побегах прошлого года, волчковые и порослевые — из спящих почек на многолетних органах. На основных побегах в пазухах их листьев развиваются пасынковые побеги, на которых могут развиваться пасынки следующих порядков. Сочетание скелетной части, основных побегов и пасынков составляет форму куста (рис. 7).

Образование стебля у сеянцев начинается при развитии первого побега из зародышевой почечки, расположенной выше корневой шейки. Формирование стебля у саженцев начинается на материнских растениях при образовании побегов, используемых для размножения, и продолжается при развитии почки зимующего глазка — стебель у них начинается от пятки подземного ствола (штамба). Размеры стебля и куста винограда зависят от биологических особенностей сорта, почвенного плодородия, климата, технологии возделывания и других факторов.
Побеги винограда состоят из узлов и междоузлий (рис. 8).

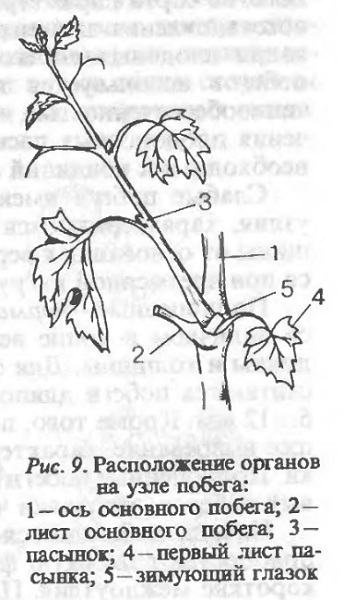
На узлах побегов текущего года развивается ряд важных вегетативных и генеративных органов виноградного растения. Листья прикрепляются к узлам посредством длинных черешков. По длине побега листья располагаются на узлах, супротивно чередуясь вдоль оси побега. У основания черешка, в месте их прикрепления к побегу, на растущей верхушке и близко к ней располагаются пленчатые прилистники, которые затем отсыхают и опадают. В пазухе листа закладываются пазушные почки (рис. 9). На нижних 2…6 узлах побегов формируются только листья и пазушные почки. На последующих узлах напротив листьев в определённой закономерности развиваются усики или соцветия. На бесплодных побегах напротив листьев образуются только усики, причём после двух узлов с усиком (симподиальный тип роста) следует узел без усика (моноподиальный тип роста).
На побегах вида V. labrusca усики образуются на всех узлах побега, кроме нижних 2…5. На самых нижних симподиальных узлах плодоносных побегов развиваются вместо усиков соцветия. На одном плодоносном побеге может развиваться от 1 до 7 соцветий, чаще 1…3. Если на побеге развился усик, то выше него соцветие сформироваться не может.
Иногда, как исключение вместо усика развивается внепазушный побег, который, подобно обычному побегу, состоит из узлов и междоузлий, несёт на себе листья, пазушные почки, пасынки усики (редко и соцветия). Пасынки по морфологии подобны основным побегам: несут на себе листья, пасынки следующего порядка, зимующие глазки, усики, иногда и соцветия.
Побеги винограда имеют различную силу роста. У жирующих побегов толщина и длина превышают средние для сорта показатели. У них удлинённые междоузлия, мелкие размеры зимующих глазков, рыхлая древесина, сильно развитая сердцевина, они слабо прикреплены к несущим элементам куста. Жирующими чаще являются волчковые и порослевые побеги. Много жирующих побегов образуется при повреждении куста, неправильной (слабой) нагрузке побегами и др. Жирующие побеги или удаляют в ранний период развития, или проводят раннюю чеканку, что позволяет получать на них несколько пасынков с типичными для побегов сорта параметрами. Их используют в последующем для восстановления элементов куста, заготовки черенков, формирования плодовых побегов и сучков. Ранняя чеканка жирующих побегов используется также в некоторых районах с высокой теплообеспеченностью и длинным периодом вегетации для получения плодоносных пасынков. Грозди для них успевают достичь необходимых кондиций созревания.
Слабые побеги имеют небольшую длину, короткие междоузлия, характеризуются быстрым и резким уменьшением толщины от основания к верхушке. Много слабых побегов образуется при чрезмерной нагрузке кустов.
Полноценные — нормально развитые — побеги характеризуются наличием в конце вегетации средних для сорта размеров длины и толщины. Для сортов вида V. vinifera L. полноценными считаются побеги длиной 100…150 см и толщиной у основания 6…12 мм. Кроме того, полноценные побеги должны иметь хорошее вызревание, характерную для сорта окраску, развитые глазки. Полноценные побеги используются для формирования плодовых побегов, заготовки черенков.
Иногда наблюдается фасциация (уродливость) побегов: они приобретают плоскую фоорму, верхушка раздваивается, образуя короткие междоузлия. Причиной этого чаще являются вирусные заболевания, повреждения гербицидами и др.
Рост стебля сеянца начинается с развития зародышевой почечки и образования первого его побега (рис. 10). При прорастании в семени активизируются процессы метаболизма, в который интенсивно вовлекаются запасные питательные вещества эндосперма, начинается деятельность меристематических тканей. Покровы семян растрескиваются, и появляется главный корень. Затем происходит рост подсемядольного колена (гипокотиля), оно приобретает изогнутую форму, а затем выпрямляется, в результате чего семядоли выносятся на поверхность субстрата. На свету семядольные листья приобретают зелёную окраску и начинают выполнять функции листа. Одновременно с развитием подсемядольного колена, представляющего собой зачаток стебля винограда, формируется первичная проводящая система, стебель начинает выполнять свои основные функции и начинает развитие зародышевой почечки, расположенной между семядолями (рис. 10).В клетках точки роста почечки возрастают процессы метаболизма, они приобретают меристематическую активность и формируют конус нарастания, при развитии которого побег растёт. По мере роста сбоку от точки роста последовательно отчленяются зачаточные бугорки листьев. В пазухах зачатков листьев по мере их развития закладываются пазушные почки. Из листового бугорка формируется листовой черешок, листовая пластинка и два пленчатых прилистника, расположенных у основания черешка с обеих сторон. Прилистники прикрывают нежную верхушку побега, в последующем по мере удаления узла, их несущего, от верхушки, отсыхают и отпадают, от них по обеим сторонам основания черенка на побеге остаются два полулунных следа.

Первый настоящий лист сеянца образуется на 22…27-й день после начала прорастания семени. В последующем, если нет повреждений, продолжается рост главного стебля (первого побега) сеянца и до конца первой вегетации на нём образуется 15…30 листьев, которые располагаются спирально по формуле 2/5. До 6…10-го узла рост побега сеянца происходит моноподиально — по мере развития центральной оси конуса нарастания сбоку отчленяются зачатки листьев, в их пазухе закладываются пазушные почки. На 6…10-м узле сбоку от главной оси в пазухе зачатка верхнего листа образуется меристиматический бугорок, который быстро развивается, обгоняет главную ось и продолжает рост побега. Главная ось замедляется в развитии, отклоняется в сторону и формирует усик. Такой рост побегов называется симподиальным.
Новая точка роста продолжает рост главного стебля до следующего узла, на котором вновь откланяется в сторону и формирует второй усик, а образовавшийся сбоку новый бугорок формирует следующую точку роста и продолжает рост побега. На третьем узле смещение точки роста не происходит и усик не образуется. В последующем по всей длине побега винограда наблюдается смешанный моноподиально-симподиальный тип роста: за каждыми двумя узлами с усиком следует один узел без усика. Это характерно для всех видов рода Vitis, кроме V. labrusca L., у которого после после появления первого усика они образуются на всех последующих узлах (рис. 11). В конце вегетации рост побега замедляется, прекращается и верхушка отмирает.

Стебель саженца растёт и ежегодно возобновляется рост у растений семенного и вегетативного происхождения в результате развития пазушных перезимовавших почек. У таких побегов первые 2…5 узлов имеют моноподиальный, а в последующем — смешанный моноподиально-симподиальный тип роста (за исключением V. labrusca).
Молодые растущие побеги текущего года травянистые, сочные, хрупкие, имеют зелёную окраску, верхушка их изогнута и имеет характерные для данного ампелографического сорта опушение и окраску. Во второй половине вегетации происходит одревеснение побегов, начиная от основания, они приобретают коричневую окраску. Это связано с формированием защитных тканей, одновременно накапливаются запасные и защитные вещества для обеспечения благополучной перезимовки.
Для виноградного побега характерна дорзивентральность (поперечная полярность), следствием которой является выраженная асимметричность его (рис. 12). Поперечный срез побега винограда имеет вид прямоугольника с неравномерно закруглёнными сторонами. Более широкие стороны — плоская (расположен усик) и желобчатая (расположен лист и пасынок). Две другие стороны — спинная и брюшная — округлены и более утолщены (особенно последняя). Зимующий глазок смещён к брюшной части, а пасынок — к спинной.

Элементы первичной анатомической структуры стебля (рис. 13) формируются в результате деления и дифференциации клеток меристемы конуса нарастания. Наружный слой клеток конуса нарастания (туника) формирует эпидермис стебля, внутренние слои (корпус) — паренхиму первичной коры и центрального цилиндра. Клетки, расположенные в основании конуса нарастания, отчленяют бугорки листьев и почек. Эпидермис побега состоит из плотно прилегающих клеток, содержащих значительное количество танина. С внешней стороны эпидермис покрыт кутикулой. Наружные эпидермальные клетки молодых побегов образуют выросты (волоски, жемчужные желёзки), а также устьица, через которые осуществляется газообмен, транспирация. Первичная кора состоит из 8…10 слоёв коровой паренхимы, слаборазвитой первичной механической ткани — колленхимы и однослойной эндодермы. Клетки коровой паренхимы имеют округлую форму и содержат углеводы, танин и большое количество хлоропластов.
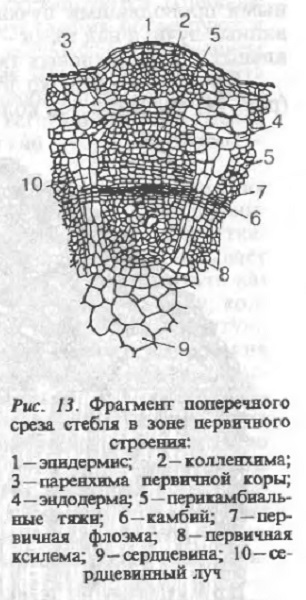
В состав центрального цилиндра входит несколько проводящих пучков (чаще 5), расположенных радиально, и сердцевины, состоящей из тонкостенных паренхимных клеток. Между первичными проводящими пучками располагаются первичные сердцевинные лучи, а над ними — механическая ткань в виде перикамбиальных и перецикловых тяжей.
Перед переходом ко вторичному анатомическому строению (рис. 14) формируется кольцо камбия. Из клеток камбия между первичным лубом и древесиной образуется пучковый камбий. Меристематическая активность возобновляется, и в перикамбиальном слое сердцевидных лучей формируется межпучковый камбий, при соединении которого с пучковым образуется сплошное камбиальное кольцо. Это происходит сравнительно близко к верхушке побега. В процессе меристиматической активности пучкового камбия образуются наружу вторичный луб (флоэма), а втутрь — вторичная древесина (ксилема). Таким образом стебель переходит ко вторичному строению. Камбий функционирует в течение всей жизни стебля, возобновляя свою активность весной и завершая осенью. В результате формируются годичные кольца. При делении клеток межпучкового камбия формируются вторичные сердцевинные лучи, которые являются продолжением первичных. Интенсивность работы камбия зависит от вида, сорта винограда, фазы вегетации, внешних условий, ухода за растением и других факторов. В первой половине вегетации формируются более крупные проводящие сосуды, мягкий луб, а во второй — образуется преимущественно либриформ.

В период замедления роста побегов и деятельности камбия из клеток перицикла, паренхимных клеток флоэмы и серцевинных лучей появляется кольцо вторичной образовательной ткани — феллогена (пробкового камбия). Феллоген откладывает наружу стебля пробку, а внутрь — феллодерму. В результате наружная часть, включая клетки перицикла, первичную кору, колленхиму, эпидермис и др., отмирает и приобретает жёлтую, а затем и коричневую окраску, начинается процесс вызревания побегов, образуется корка побега. Этот процесс начинается у основания побега и продолжается акропетально. У морозоустойчивых видов и сортов феллоген закладывается в более глубоких слоях стебля. У некоторых сортов формируется второе кольцо феллогена под первым слоем твёрдого луба, в результате чего образуется двухслойная перидерма. В последующие годы феллоген образуется в более глубоких слоях флоэмы, что приводит к образованию комплекса отмерших защитных тканей — корки. В результате утолщения стебля наружные слои корки растрескиваются.
Одновременно с вызреванием побегов наружные слои клеток сердцевины, прилегающие к древесине, опробковевают. В результате клетки сердцевины отмирают и наполняются воздухом — формируется аэренхима, которая облегчает массу стебля при прикреплении к опоре и улучшает возможности для газообмена. На узлах аэренхима прерывается диафрагмой — особой тканью, состоящей из живых крупных, плотно прилегающих друг к другу клеток, содержащих большие запасы питательных веществ, особенно крахмала. Диафрагма на узлах с усиком полная и неполная — на узлах без усика.
Побег растёт за счёт деления, растяжения и дифференциации клеток в результате жизнедеятельности различных меристем. Апикальная меристема обеспечивает рост в длину, образование других надземных органов. Утолщение побега происходит в результате функционирования камбия, а вызревание — феллогена. Интеркалярная меристема образуется от апикальной. За счёт интеркалярного удлиняется побег. Максимальное растяжение наблюдается у 2…3-го сверху междоузлия, затем замедляется и прекращается, когда оно становится 5…7-м. Таким образом, зона роста побега значительно длиннее, чем такая же у корня. Количество растущих междоузлий зависит от роста: чем интенсивнее рост, тем больше междоузлий. Рост побегов начинается весной после распускания почек, по мере повышения температуры интенсивность усиливается и достигает максимума в период, близкий к цветению, затем постепенно замедляется и прекращается. Для побега винограда характерна очень высокая интенсивность роста: суточный прирост составляет в период максимальной интенсивности 4…5 см, иногда 10 см и более. Динамика интенсивности роста пасынков так же, как у основных побегов, представляет собой одновершинную кривую.
В период роста верхушка побега изогнута, что связано с неравномерностью роста на разных сторонах побега — на спинной стороне рост клеток происходит интенсивнее. При наклонном или горизонтальном положении растущего побега его брюшная сторона направлена к поверхности земли, а желобчатая и плоская чередуются.
На рост побегов влияет ряд внутренних и внешних факторов. С повышением роста корней усиливается рост побегов. Чем мощнее корневая система, тем больше побегов развивается на кусте, при этом недостаток точек роста из-за сильной обрезки или повреждение надземной части куста компенсируется развитием спящих почек. Сила роста побегов зависит от их количества на кусте: с увеличением числа побегов сила роста каждого падает. Существует корреляция между растущей верхушкой побега и развитием пасынков. Весной пасынковые почки и побеги развиваются слабо, если растёт основной побег, их несущий. При повреждении точки роста основного побега стимулируется развитием пасынковых почек и рост пасынков. Причём сила роста пасынков повышается по мере приближения их к верхушке основного побега, а самый верхний из них приобретает максимальную силу роста и заменяет основной побег. Коррелятивные взаимоотношения проявляются не только при повреждении верхушки, но и при изменении угла наклона побега по отношению к поверхности почвы. Вертикальное положение побега тормозит рост пасынков и, наоборот, ориентация его наклонно, горизонтально или верхушкой вниз стимулирует развитие боковых органов (пасынки, пазушные почки, генеративные органы). Существует связь между отдельными побегами куста: сильное развитие одних замедляет рост других. Поэтому задача состоит в том, чтобы разместить многолетние части, плодовые и растущие побеги таким образом, чтобы обеспечить равномерность роста побегов, стимулировать рост тех из них, которые необходимы для формирования плодовой древесины в будущем году.
Развитие глазков, сила роста побегов из них зависят от положения плодового побега в пространстве. Вертикальное и близкое к нему положение плодового побега стимулирует развитие верхних глазков и тормозит развитие нижних, а при наклонном и горизонтальном положении его обеспечивается равномерное развитие глазков и одинаковая сила роста побегов. При изгибе плодового побега в виде дуги или кольца наблюдается усиленный рост побегов и глазков, расположенных ближе к основанию, и ослабление верхних. Установлена корреляция, заключающаяся в том, что на той стороне куста, где сильнее развиты корни, также сильнее развиты побеги.
Важнейшим внутренним фактором, регулирующим рост побегов, являются гормоны. Баланс гормонов различной природы, их взаимодействие определяет не только силу роста побегов, но и ход других важнейших физиологических процессов. Путём экзогенного применения регуляторов роста можно изменить гормональный статус растения или его органа и получить необходимый эффект. Установлено, что ауксины задерживают рост боковых почек, стимулируют рост корней. Гиббереллины стимулируют рост побегов и плодов, но задерживают развитие корней. Ретарданты тормозят рост побегов, стимулируют развитие боковых органов и т. д.
На процессы роста побегов влияют сорт, внешние условия, агротехника. Разным сортам свойственна различная сила роста побегов, поэтому существует классификация сортов по силе роста: сильно-, средне- и слаборослые. Из внешних условий важное значение имеют температура, влага, питание (особенно азот).
На силу роста пасынков влияют сортовые особенности, почвенно-климатические условия, уход, нагрузка куста, рост основного побега и др. Сильно растут они на сортах, склонных к сильному росту пасынков, на плодородных почвах, при благоприятном водном режиме. Усиленный рост пасынков отмечается при нарушении корреляции между корневой системой надземной частью, низкой нагрузке куста побегами, горизонтальном или свободном свисании основных побегов. При повреждении верхушки или её удалении у основного побега рост пасынков усиливается, особенно самого верхнего. Пасынки имеют такое же строение, как и основные побеги. Усиленный рост их приводит к ослаблению роста основного побега, загущению кроны куста. Для предотвращения этого производят пасынкование. Особенно это необходимо для маточников, где усиленный рост пасынков снижает выход и качество черенков.
Усики на пасынке появляются, как правило, со второго узла. При благоприятных для развития пасынка условиях на нём могут образовываться соцветия. Это свойственно используют для получения дополнительного урожая на пасынках при низком основном урожае, повреждении кустов. Для этого проводят раннюю прищипку жирующих основных, чаще бесплодных, побегов. Такой приём сочетают с усилением ухода за кустами. В результате образуются сильно растущие плодоносные пасынки, урожай которых при благоприятных климатических условиях (высокая сумма активных температур, длинный период вегетации, сильнорослые сорта) достигает необходимых кондиций, хотя и позже основного урожая. Сильные, хорошо вызревшие пасынки используют также для быстрого восстановления структуры куста. Такой приём на жирующих побегах маточных кустов позволяет получать пасынки, соответствующие параметрам полноценного побега и пригодные для заготовки черенков.





