К отделу относятся облигатные внутриклеточные паразиты.
Вегетативное тело в виде многоядерного протопласта, не способного к самостоятельному движению и находящегося внутри клетки растения-хозяина.
Специальные спороношения не образуются. Зимующая стадия представлена спорами, образующимися при распаде внутриклеточного плазмодия. При наступлении благоприятных условий споры прорастают двужгутиковыми зооспорами. Жгутики гладкие, прикреплены к переднему концу, один из них длинный, другой – короткий.
В жизненном цикле имеется две стадии: гаплоидная и диплоидная.
Пораженные ткани разрастаются и формируют наросты неправильной формы на подземных органах растений.
Классификация. Отдел включает один класс Plasmodiophoromycetes – плазмодиофоромицеты (1 порядок, 1 семейство, 30 видов). Распространенные роды: Plasmodiophora (плазмодиофора), Spongospora (спонгоспора). Всего 30 видов.
Класс Plasmodiophoromycetes – плазмодиофоромицеты. Порядок Plasmodiophorales – плазмодиофоральные. Семейство Plasmodiophoraceae – плазмодиофоровые (Plasmodiophora, Spongospora).
Род Plasmodiophora – плазмодиофора.
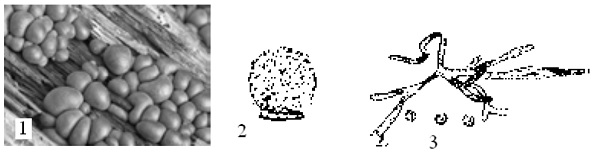
Наиболее известный представитель – Plasmodiophora brassicae (плазмодиофора капустная), поражающий корни растений из семейства крестоцветные. Заболевание известно под названием «кила капусты». Пораженные корни растения имеют уродливый вид, который придают им объемные опухоли (рис.10, 1). Плазмодий паразита находится внутри клеток корня растения (рис. 10, 2). Под влиянием паразита нарушается синтез фенольных и индольных соединений в клетках растений, что приводит к увеличению размеров клеток (гипертрофии) и усиленному их делению (гиперплазии). Впоследствии пораженные корни загнивают. В клетках корня находятся многочисленные споры (рис. 10, 3). При заражении капустной рассады кочаны могут совсем не развиваться. У зараженных взрослых растений они недоразвиваются.
Рис. 10. Plasmodiophora brassicae – плазмодиофора капустная: 1 – внешний вид корней пораженного растения; 2 – клетки растения с плазмодием паразита; 3 – клетки растения с гаплоидными спорами паразита.
В цикле развития различают несколько стадий, протекающих в корневых волосках, почве и клетках коры корня растения (рис. 11).
Первичная стадия. Протекает в почве и корневом волоске.
Покоящиеся гаплоидные споры (рис. 11, 1), находящиеся в почве, прорастают первичными зооспорами (рис. 11, 2), имеющими два гладких жгутика разной длины на апикальном конце.
В корневом волоске (n).
Зооспоры достигают корневого волоска, теряют жгутики и прикрепляются к нему с помощью выроста в виде трубки. В ней находится «шип» из плотной плазмы, который прокалывает оболочку корневого волоска. Плотное прижатие «шипа» к поверхности волоска обусловлено образованием в бывшей зооспоре вакуоли. Цитоплазма переливается в корневой волосок через тонкий канал, ядра митотически делятся с образованием гаплоидного первичного плазмодия (рис. 11, 3 – 5). Затем плазмодий распадается на одноядерные фрагменты округлой формы (рис. 11, 6). Ядра в них делятся несколько раз митотически (рис. 11, 7). Содержимое распадается на 4 – 8 частей (по числу ядер), которые становятся вторичными зооспорами (или гаметами), выходящими из корневого волоска в почву (рис. 11, 8).
Вторичная стадия. Протекает в почве и клетках коры корня.
В почве. Зооспоры, функционирующие как гаметы, попарно сливаются (плазмогамия), образуется двуядерная клетка, которая способна заражать корни растения (рис. 11, 9 – 10).
В клетках коры корня. В корнях растения ядра митотически делятся, развивается сначала двуядерный вторичный плазмодий (рис. 11, 11), а затем мощный вторичный многоядерный плазмодий (рис. 11, 12 – 13). Пораженные корни утолщаются. На поздних стадиях развития болезни ядра плазмодия попарно сливаются (кариогамия), далее следует редукционное деление (рис. 11, 14), и плазмодий распадается на массу гаплоидных мелких круглых спор (мейоспоры) (рис. 11, 15).
В почве. Споры при сгнивании корней попадают в почву. Их распространению в почве способствуют животные (дождевые черви, насекомые), токи воды (дождь, полив), деятельность человека, например, перенос земли со спорами на орудиях обработки почвы, и т. п. В почве споры могут годами сохраняться, не теряя способности к прорастанию. При благоприятных условиях и стимулирующем действии корневых выделений растений-хозяев споры прорастают. Впервые капустную килу тщательно изучил М. С. Воронин (1878). Он установил причину заболевания, описал в главных чертах цикл развития паразита, предложил меры борьбы с ним, а также указал на его способность заражать другие растения из семейства крестоцветных. Этот гриб может поражать около 200 видов культурных и дикорастущих растений из сем. крестоцветные. Кислая реакция почвы способствует заражению, поэтому в борьбе с этим заболеванием производят её известкование.
Рис. 11. Схема цикла развития Plasmodiophora brassicae (пояснения в тексте).
Род Spongospora – спонгоспора.
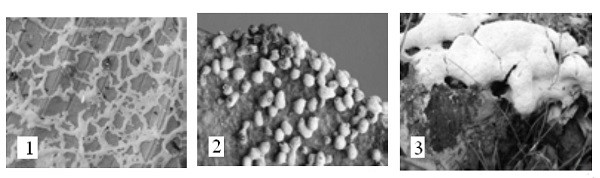
Spongospora solani – спонгоспора картофельная. Является возбудителем порошистой парши клубней картофеля (рис. 12). Многоядерные плазмодии спонгоспоры развиваются в периферических клетках клубня. Затем плазмодий распадается на споры, плотно склеивающиеся в губчатые комочки. Пораженный клубень покрыт язвочками с коричневым порошистым содержимым, представленным скоплением этих комочков. Цикл развития похож на цикл развития Plasmodiophora brassicae. Иногда поражаются стебли и столоны. Кроме картофеля поражаются томаты и другие растения сем. пасленовые.
Рис. 12. Spongospora solani – спонгоспора картофельная, возбудитель порошистой парши картофеля: 1 – внешний вид пораженного клубня; 2 – комочки из спор в клетке клубня.
Название «слизевики» связано с обликом вегетативного тела, представленного слизистым голым многоядерным протопластом – плазмодием, размеры которого могут быть микроскопическими либо макроскопическими, достигающими несколько десятков сантиметров в диаметре. В состав плазмодия входит вода (до 75%), белки (около 30%), гликоген, пигменты, придающие яркую (желтую, красную, фиолетовую и др.) окраску. Слизевикам с окрашенными плазмодиями необходим свет для нормального спороношения, так как некоторые пигменты играют роль фоторецепторов. В плазмодиях обнаруживают пульсирующие вакуоли. Плазмодий способен к активному передвижению (переползанию) к источнику пищи, воды, света.
Вегетативная стадия сменяется различными формами спороношений. Наиболее простой тип: весь плазмодий покрывается оболочкой и приобретает форму лепешки, подушечки. В других случаях образуются более сложные спороношения (шаровидные, цилиндрические, ветвистые и др.). Они могут быть сидячими или на ножках. Спорангий покрыт оболочкой, в ряде случаев содержащей целлюлозу и известь. Формирующиеся спороношения могут сливаться, образуя эталий, имеющий общую наружную оболочку. Внутри спороношений развиваются споры с многослойной оболочкой. Поверхность спор часто неровная: с шипиками, бородавками и т.п. У многих слизевиков внутри спороношения образуются из вытягивающихся и сливающихся вакуолей гигроскопические переплетающиеся полые или сплошные нити – капиллиций. При изменении влажности капиллиций способствует разрыхлению споровой массы и распространению спор. В эталиях эту же функцию выполняет псевдокапиллиций, состоящий из остатков слившихся спорангиев. Перед образованием спор идет редукционное деление.
Гаплоидные споры могут прорастать в воде или на влажном субстрате. В первом случае из споры образуется 1–8 зооспор с одним или двумя гладкими неравными апикальными жгутиками. В отсутствие воды спора прорастает безжгутиковыми миксамёбами, которые перемещаются с помощью выдвигающихся и втягивающихся псевдоподий. И зооспоры, и миксамёбы могут попарно сливаться с образованием диплоидной клетки. Это явление считается половым процессом у миксомицетов. Далее в результате митотических делений увеличивается количество ядер и развивается диплоидный плазмодий.
В циклах развития преобладает диплоидная стадия (диплонты).
Питание происходит путем внутреннего переваривания частиц пищи. Источником питания могут быть бактерии, клетки водорослей, гифы грибов.
Большинство миксомицетов являются космополитами. В основном это сапротрофы, обитающие в почве, на лесной подстилке и растительных остатках в лесных экосистемах. Реже их можно обнаружить на остатках животного происхождения. Наиболее активно споры образуются в летне-осенний период. Для развития плазмодия необходима высокая влажность субстрата, а для образования спор – более низкая. Поэтому миксомицеты чаще всего встречаются в лесах умеренного климата, чем в тропических.
Доказано, что по происхождению это – полифилетическая группа.
Классификация. В отделе выделяют два класса – Myxomycetes (миксомицеты) и Protosteliomycetes (протостелиомицеты).
В основе выделения порядков лежат особенности строения спорангия.
Типы спороношений
Плазмодиокарп. Наиболее простой тип спорофора. Весь плазмодий покрывается оболочкой и превращается в спорофор. Поэтому они всегда сидячие.
Спорангий. Плазмодий распадается на множество образований, формирующих дифференцированные спорангии, сидячие или состоящие из головки с ножкой.
Эталии образуются из ветвящихся плазмодиев. Спорангии сливаются, покрываются общей оболочкой.
Псевдоэталии. Спорангии плотно прилегают друг к другу, но боковые стенки сохраняются, поэтому спорангии не теряют индивидуальности. Они могут быть сидячими и на ножках.
Между всеми типами спороношений имеются переходы.
Порядок Liceales – лицеальные
Характерно наличие спороношений в виде спорангиев, эталиев. Настоящий капиллиций отсутствует, имеется псевдокапиллиций. Споры светлые или ярко окрашенные. Род Lycogala (ликогала). Lycogala epidendrum (ликогала древесинная). Народное название гриба – «волчье вымя». Встречается повсюду на пнях, древесных остатках. Плазмодий ярко окрашен, кораллово-красного цвета. Спороношение в виде эталиев с гладкой оболочкой красно-розового цвета. Содержимое эталиев такого же цвета, слизистое. По мере созревания эталии становятся буровато-серыми. Наверху образуется отверстие, через которое высыпаются споры, что напоминает маленькие дождевики (около 1 см в диаметре) (рис. 6).
Рис. 6. Lycogala epidendrum – ликогала древесинная: 1 – эталии; 2 – зрелый эталий в разрезе; 3 – нити псевдокапиллиция со спорами.
Порядок Physarales – физаральные
К этому порядку относится наибольшее число видов. Спороношения в виде сидячих или на ножке спорангиев, эталиев. Развит капиллиций в виде трубочек различной формы. На поверхности оболочек спорангия, спор и капиллиция откладывается известь. Споры темные. Род Physarum – физарум. Наиболее представлен по количеству видов. Широко распространен Physarum cinereum (физарум пепельный), встречающийся на опаде, иногда даже на живых травянистых растениях, растущих на газонах. Может вызывать угнетение растений. Плазмодий сначала белый, затем желтеет. Спорангии сидячие, перидий пепельно-серого цвета (рис. 7). Споры темные, почти черные. Группа спорангиев может покрываться общей оболочкой, не теряя своей индивидуальности, с образованием плазмодиокарпа.
Род Fuligo – фулиго. На территории России широко распространен Fuligo septica (фулиго септированный). Встречается на гниющих растительных остатках. Плазмодий желтого, бежевого, беловатого цвета. Спороношение чаще всего в виде сидячих эталиев, более 1 см диаметром. Перидий содержит известь, поэтому очень хрупкий. Развит псевдокапиллиций (тоже с зернами извести). Споры темные, грязно-черные.
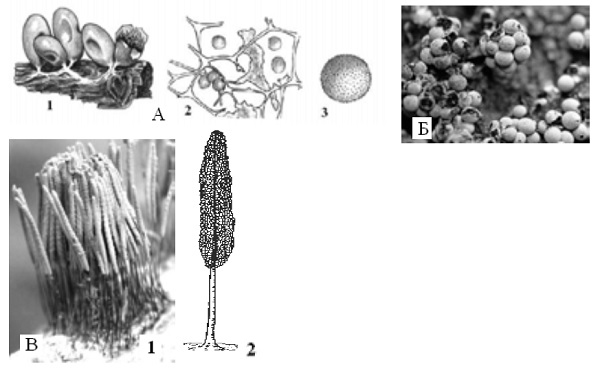
Род Leocarpus – леокарпус. Довольно часто встречается на мхах и живых травянистых растениях, на гнилых растительных остатках леокарпус хрупкий (Leocarpus fragilis). Его блестящие, коричневого цвета спорангии содержат темные споры. Перидий спорангия трехслойный, хрупкий. Наружный и внутрениий слои – тонкие, а средний, обызвествленный, – толстый. Колонка отсутствует. Капиллиций состоит из плоских, расширенных в местах ветвления трубочек без извести (рис. 8, А). Род Didymium – дидимиум (рис. 8, Б). Спорангии сидячие или на ножке, могут быть плазмодиокарпы. Споры черные. Перидий однослойный или двухслойный с включениями кристаллической извести. Капиллиций без извести. Встречается на гнилых растительных остатках, опаде, травянистых растениях. Спороношение в виде отдельных спорангиев (часто на ножке) или эталиев без извести. Развит капиллиций, также не содержащий извести. Споры темные. Род Stemonitis – стемонитис. Плазмодий тонкий, прозрачный. Спорангии вытянутые, на ножке, которая образует внутри них колонку. Перидий быстро исчезает, обнажается капиллиций (без извести), похожий на ажурные перышки коричневого цвета. Встречается на гнилой древесине и других отмерших растительных остатках (рис. 8, В).
Рис. 8. A. Leocarpus fragilis – леокарпус хрупкий: 1 – спорангии; 2 – капиллиций со спорами; 3 – спора. Б. Didymium squamulosum – дидимиум чешуйчатый (спорангии). В. Stemonitis fusca – стемонитис бурый: 1 – внешний вид спороношения; 2 – капиллиций.
Порядок Trichiales – трихиальные
Спорангии светлые, сидячие или на ножке. Развит капиллиций различного строения: нити со спиральными, кольчатыми утолщениями или без них, ветвящиеся или не ветвящиеся. Споры светлые, желтоватые. Род Trichia – трихия (рис. 9). Представителей рода можно обнаружить на гнилой древесине, коре живых деревьев. Чаще всего образуются спорангии, близко расположенные друг к другу. Они могут быть сидячими или на ножке.
Рис. 9. Trichia varia – трихия изменчивая: 1 – группа спорангиев; 2 – вскрывающиеся спорангии; 3 – нити капиллиция со спиральными утолщениями.
Еще в конце XIX в. А. де Бари (1884) относил миксомицеты к царству животных, поскольку эти организмы обладают чертами животных и грибов. С животными их сближают зоотрофное питание, наличие вегетативной амебоидной стадии, присутствие в циклах развития подвижных стадий (с двумя апикальными гладкими жгутиками), строение митохондрий. В настоящее время во многих системах органического мира они включены в царство Protozoa. Из всего многообразия отделов охарактеризуем только два – Myxomycota и Plasmodiophoromycota.
Грибы появились на нашей планете около 1,3 млрд. лет назад в протерозойском эоне. В строматолитах (кораллоподобные осадочные образования, карбонатные или кремниевые) этого периода обнаружены грибоподобные организмы, которые были похожи на дрожжи и мукоровые грибы. В целом палеонтологические свидетельства разнообразия грибов немногочисленны. Более устойчивы к воздействиям внешней среды споры грибов, поэтому спорово-пыльцевой анализ широко используется в палеомикологии. В фанерозойский эон (палеозойская эра), в верхнем докембрии, обитали водные грибы. В кембрии (около 600 млн. лет назад) появляются хитридиевые грибы, а уже в силуре (около 430 млн. лет назад) наблюдается переход грибов от водного образа жизни к наземному (рис.5). В это время на растительных остатках в прибрежных зонах обитали «слизистые» оомикота, аско- и дейтеромикота.
Рис. 5. Время (млн. лет) возникновения основных таксономических групп грибов
В девоне (около 400 млн. лет назад) эндофитные грибы рода Glomites обнаружены в окременелых тканях осевых органов риний. Наряду с симбиотическими у них были выражены патогенные свойства. Эндомикориза была широко распространена в раннем девоне. К этому времени относятся находки первых древних лишайников (цианолишайники). В девоне также получают развитие паразитические грибы и сапротрофы (аскомикота). Сумчатые грибы, таким образом, являются более древними в сравнении с базидиальными. Данные молекулярной генетики (анализ последовательностей 18 SPHK с использованием базы данных банка генов, число мутационных замен в ряде генов) подтвердили палеонтологические свидетельства о том, что дивергенция грибов произошла около 400 млн. лет назад, когда появились наземные растения. В конце палеозойской эры (пермский период, 280 млн. лет назад) палеонтологические находки говорят о наличии «белой гнили» растений, вызываемой обычно базидиальными грибами. В мезозойскую эру (триасовый период, 225 млн. лет назад) параллельно с развитием голосеменных растений возникает эктомикориза, образуемая агариковыми грибами. Основное развитие микоризы приходится на меловой период (около 130 млн. лет назад). В это же время обнаружены плодовые тела агариковых грибов. Имеются достоверные находки окремненных остатков агарикового гриба археомарасмиус (Archaeomarasmius legetti – 90–94 млн. лет назад), похожего на современных представителей рода Marasmius (негниючник). Таким образом, грибы являются одними из древнейших организмов на нашей планете. Уже в палеозойскую эру (девон, около 400 млн. лет назад) появились основные группы грибов, трофически и топически связанных с растениями и животными. Их эволюция шла в основном совместно с эволюцией растений (коэволюция). Грибы способствовали освоению растениями суши, их процветанию и развитию, что привело к многообразию растений и наземных экосистем.
Гипотезы происхождения грибов
Вопрос о происхождении грибов до сих пор окончательно не решён. Предполагают, что грибы возникли несколькими самостоятельными эволюционными линиями. К настоящему времени известно несколько гипотез происхождения грибов.
Происхождение от каких-то бесхлорофилльных амебообразных или жгутиконосных эукариот – зоофлагеллят. В результате приспособления к наземному образу жизни жгутики у многих видов были утрачены.
Предками некоторых отделов грибов являются водоросли (цианобактерии, бурые, красные и др.). Возможно, сумчатые грибы (а от них – базидиальные) произошли от безжгутиковых красных водорослей из класса Florideophyceae. У красных водорослей есть общие черты с сумчатыми грибами: строение пор – септ (перегородок между клетками), в оболочках некоторых водорослей обнаружен хитин, имеются сходные по химическому составу метаболиты. У багрянок во время митоза, как и у грибов, сохраняется ядерная оболочка (закрытый митоз). Достаточно много общего у красных водорослей и цианобактерий (прокариоты), в частности, состав пигментов и некоторых других соединений. Поэтому согласно другой гипотезе (парафлоридейная гипотеза) сумчатые грибы и багрянки произошли от общего предка, но затем развивались независимо друг от друга. Оомикота являются самостоятельной эволюционной линией, берущей начало от форм, близких к золотистым или желто-зеленым водорослям.
Происхождение от прокариотного организма, близкого к Eubacteria. Фотосинтезирующая бактериальная клетка в результате совершенствования структур дала начало простейшей эукариотной клетке. Эволюция шла через промежуточный дрожжеподобный организм протомицес (Protomyces), давший начало развитию грибной клетки. Доказательством тому является примитивное строение клеток аскомицетных дрожжей: в них отсутствует аппарат Гольджи, а также пульсирующие вакуоли и жгутики. Митохондрии примитивные, геном минимальный (сравним лишь с геномом цианобактерий), химический состав клеточных стенок близок к прокариотам. Эта гипотеза подтверждается современными данными об ультраструктурном строении клеток прокариот и эукариот.
На основании изучения последовательностей малых субъединиц рибосомальной РНК в 1993 г. было высказано предположение о том, что предком животных и грибов был единый прокариотный жгутиковый организм. Принимая во внимание биохимические критерии, следует считать, что им являлись не пурпурные бактерии, а актиномицеты.
Практически все живые организмы можно разделить по структурно-морфологическому строению на две группы (надцарства, империи): прокариоты (доядерные) и эукариоты (ядерные). Прокариоты (Procaryota) не имеют ядерной оболочки, хромосом. Настоящий половой процесс (мейоз) и митоз отсутствуют. Нет пластид, митохондрий, эндоплазматической сети, телец Гольджи, микротрубочек. В клеточной стенке может быть глюкопептид муреин. Жгутиков чаще всего нет или они просто устроены. Пищевых вакуолей нет, но имеются газовые вакуоли. Многие могут фиксировать атмосферный азот, дыхание анаэробное и аэробное, чувствительны к антибиотикам, но устойчивы к облучению. К прокариотам относятся бактерии, актиномицеты, цианобактерии (сине-зеленые водоросли). Все остальные организмы относятся к эукариотам. Эукариоты (Eucaryota) имеют ядро, окруженное мембраной, генный материал находится в хромосомах, состоящих из нитей ДНК и белков. Имеются настоящий половой процесс, мейоз и митоз. В клетках могут быть пластиды, митохондрии, эндоплазматическая сеть, тельца Гольджи, микротрубочки, пищевые вакуоли. Жгутики сложного строения. Как правило, эукариоты не фиксируют атмосферный азот, являются аэробами или вторичными анаэробами. Не чувствительны к антибиотикам, но чувствительны к облучению. Другой принцип классификации организмов – эколого-трофический. В соответствии с ним выделяют царства: растения, животные, грибы. К растениям относятся автотрофные (фотосинтезирующие) организмы. В экосистемах они являются продуцентами. К животным относятся зоотрофные организмы, питающиеся другими организмами. Они составляют группу консументов. Для грибов характерно осмотрофное питание (поглощение веществ всем телом). Они являются редуцентами. Примером реализации изложенного подхода при построении классификационных систем являются системы Р. Уиттекера (Whittaker, 1969 – 2 надцарства, 5 царств) и А. Л. Тахтаджяна (1973, 1976 – 2 надцарства, 4 царства). Система Р. Уиттекера, усовершенствованная Л. Маргелис (1983), показана в упрощенном варианте на рис. 4. К прокариотам относится царство монеры – Monera. Эукариоты объединяют четыре царства: растения, животные, грибы, различающиеся способами питания (фототрофный, зоотрофный, осмотрофный), способами развития, и царство протоктисты (Protoctista), или протисты (Protista), где сосредоточены организмы простого строения, имеющие чаще всего жгутиковые стадии в цикле развития. Отражённые на схемеотделы организмов, относящиеся к протистам, ранее рассматривались в составе царства Грибы. Некоторые из них (Oomycota, Hyphochytridiomycota, Labirintulomycota) имеют мицелиальные талломы, а также другие морфологические структуры, сходные с грибами. Поэтому их называют грибоподобными протистами, грибоподобными организмами, псевдогрибами. Таким образом, грибы (в традиционном понимании) рассредоточены в двух царствах (протисты и грибы).
Рис. 4. Схема строения органического мира (упрощенный вариант)
В более поздних системах грибы входят в состав нескольких царств. Так, Т. Кавалье-Смит (Cavalier-Smith, 1998) предложил систему органического мира, состоящую из двух надцарств и шести царств. Прокариоты содержат одно царство (Bacteria), а эукариоты распределены между пятью царствами (Animalia, Protozoa, Fungi, Chromista, Plantae). Организмы, ранее относившиеся к царству Mycota (грибы – в традиционном понимании), входят в состав трех царств: Fungi (собственно грибы), Chromista (хромисты, куда включены и многие водоросли), Protozoa (простейшие животные). Разделение грибов по трем царствам основано на ряде признаков (состав клеточной стенки, путь синтеза лизина, подвижность в вегетативном состоянии, строение зооспор и гамет), что соответствует их происхождению тремя самостоятельными эволюционными линиями (более подробно эти вопросы будут освещены в соответствующих разделах).
Царство Chromista включает водоросли (бурые, желто-зеленые, золотистые, диатомовые), а также грибоподобные организмы – Oomycota, Hyphochytridiomycota, Labirintulomycota, для которых характерно наличие зооспор с двумя гетероконтными жгутиками, гладким и перистым (у гифохитридиомицетов – один перистый). В клеточной стенке чаще всего присутствует целлюлоза, синтез лизина идет по типу автотрофных организмов (у лабиринтуломицетов лизина нет), митохондрии в основном с трубчатыми кристами.
Царство Protozoa наряду с простейшими животными составляет группа «миксомицетов» (Acrasiomycota, Dictyosteliomycota, Myxomycota, Plasmodiophoromycota), имеющих вегетативную амебоидную стадию, зоотрофное питание, в циклах развития – подвижные стадии (с двумя апикальными гладкими жгутиками), митохондрии в основном с трубчатыми кристами.
Царство Fungi, Mycota образуют грибы, не имеющие подвижных стадий (исключение – хитридиомицеты с одним гладким жгутиком). Основной компонент клеточной стенки – хитин, синтез лизина идет по типу животных, митохондрии с пластинчатыми кристами, диктиосомы отсутствуют. Современные системы базируются в основном на данных молекулярной биологии, электронной микроскопии, биохимии. Количество надцарств (империй) и царств в этих системах значительно варьирует и достигает порой десятков. Особенностью указанных систем является то, что в состав даже одного царства входят растения (водоросли), животные (простейшие) и грибы. Далее приведен сокращенный вариант классификации грибов, изложенный в «Словаре грибов» 9-е изд. (2001) (www.indexfungorum.org) с некоторыми изменениями, принятыми в настоящем издании (табл. 2).
Систематика – наука о разнообразии организмов и их взаимоотношениях друг с другом. Систематика грибов является одним из основных разделов микологии. Задачи систематики – описание, определение видов и их классификация, восстановление путей эволюционного развития грибов. Системы могут быть искусственными, естественными и филогенетическими. В искусственных системах объединение видов осуществлялось на основе одного или немногих общих морфологических признаков. Такие системы не всегда отражали генетические связи между группами организмов. Венцом искусственных систем явилась система органического мира, созданная К. Линнеем. В ней грибы и некоторые другие организмы были отнесены к XXIV классу под названием «хаос». Естественные системы основаны на сходстве большого количества признаков (не только морфологических). На основе естественных создаются филогенетические системы, в которых отражаются эволюционные связи между организмами.
Грибы чрезвычайно трудны для систематики по ряду причин.
У грибов в отличие от высших растений невелико разнообразие морфологических признаков. Особенно это касается вегетативной части таллома. Имея перед собой лишь мицелий гриба трудно установить принадлежность его даже к отделу. Например, у сумчатых и базидиальных грибов мицелий с перегородками. Различие же их лишь в том, что у базидиальных грибов обычно (но не всегда) над перегородкой образуется вырост – пряжка.
Грибы часто обладают одинаковым строением в силу конвергентного сходства, обусловленного, например, паразитическим образом жизни. На схеме (рис. 2) показано, что виды А, В, С в онтогенезе имеют похожие стадии развития: D1, D 2, D 3.
Рис. 2. Конвергентное сходство грибов
Грибы имеют особый тип эволюционного процесса (симгенез). В результате этого процесса объединяются геномы организмов (А + В), относящихся даже к разным царствам. Новый организм (С) дает начало новой эволюционной линии (рис. 3).
Рис. 3. Упрощенная схема симгенеза
По грибам существует мало палеомикологических материалов. Так как споры грибов довольно устойчивы к факторам внешней среды, то спорово-пыльцевой анализ широко используется в палеомикологии. Имеются отпечатки растений с паразитическими грибами, а также окремненные остатки шляпочных и трутовых грибов.
Методы изучения грибов
На разных этапах развития систематики грибов преобладали определенные методы их изучения, что было обусловлено уровнем знаний и развитием техники.
До XX в. наиболее распространенным методом построения классификационных систем был сравнительно-морфологический.
Позднее большое значение приобретает онтогенетический подход, что особенно важно для выявления разных стадий развития одного вида в ходе онтогенеза, особенно сумчатых грибов.
В последнее время развивается ультраструктурный подход. При классификации грибов учитывается строение крист митохондрий, наличие сформированных диктиосом, строение жгутикового аппарата, порового аппарата септ.
Знание различий биохимического состава грибов применяется в хемотаксономии. При этом учитываются состав полисахаридов клеточной стенки, путь синтеза лизина и др.
С 90-х гг. XX в. при построении систем используется молекулярная систематика (геносистематика), основанная на выяснении нуклеотидной последовательности определенных генов. Методы геносистематики имеют как положительные, так и отрицательные черты. Положительным является то, что они объективны, воспроизводимы, на молекулярном уровне редко возникает конвергенция, что позволяет проверить гипотезы филогенетических связей. Отрицательной оказывается тенденция судить об эволюции организмов по результатам исследования эволюции отдельных генов (18 S и 28 S РНК).
Номенклатура и таксономические категории грибов
В названиях грибов, как и в названиях других организмов, используется бинарная номенклатура, предложенная К. Линнеем. Основной единицей в систематике является вид. Каждый вид имеет научное название, состоящее из двух слов, обозначающих родовую принадлежность и видовой эпитет. Кроме того, нередко существуют народные названия, которых у организма может быть множество, что часто приводит к путанице. Например, Leccinum scabrum (Bull.: Fr.) S.F. Gray (первое слово – род, а все вместе – вид) – обабок обыкновенный. По-русски этот гриб называется березовик, подберезовик, обабок. После названия вида следуют сокращенные фамилии авторов. В скобках указан автор, который впервые обнародовал видовое название гриба. За скобками приводится автор, который произвел изменения и предложил данную комбинацию видового и родового названия. Существует Международный кодекс ботанической номенклатуры (МКБН), в котором детально разработаны правила наименования растений и грибов.
Таксоны выше рода имеют названия с унифицированным окончанием, по которым можно определить принадлежность вида к тому или иному таксономическому рангу. Например, обабок обыкновенный (подберезовик) в системе таксонов будет выглядеть следующим образом (табл. 1).
Таблица 1
Кроме перечисленных основных таксономических рангов (вид, род, семейство, порядок, класс, отдел, царство) в сложных системах используются такие категории, как подцарство, подотдел, подкласс, подпорядок, подсемейство, подрод. У вида могут быть разновидности (Varietas), формы (Forma). При необходимости царства объединяют в надцарства (империи), отделы – в надотделы, классы – в надклассы и т. д.
Грибы – обширная группа организмов, включающая около 70–120 тыс. видов. Это лишь незначительная часть предполагаемого их количества. Так, ещё Э. М. Фриз (1794–1878) считал грибы самой многочисленной группой растительных организмов. В настоящее время Д. Хоуксворд предполагает, что существует около 1,5 миллионов видов грибов. Шляпочные грибы известны человеку с глубокой древности. В своих трудах упоминали о некоторых съедобных (шампиньоны, трюфели) и ядовитых грибах такие известные ученые, как Аристотель (IV в. до н.э.), Теофраст (III в. до н. э.), Диоскорид (I в.). Плиний-младший (I в.) обратил внимание на обилие трутовиков на стволах деревьев и причислил эти организмы к грибам. Первые попытки классификации грибов принадлежат именно ему. Он делил все грибы на съедобные и ядовитые. В Риме среди съедобных ценился цезарский гриб. Римляне были хорошо осведомлены о ядовитых свойствах грибов и умело использовали их для устранения неугодных. Предположительно, ядовитые грибы стали причиной смерти римского императора Клавдия, французского короля Карла VI, папы римского Клемента VII. Грибам поклонялись племена ацтеков, о чем говорят находки каменных статуэток грибов (рис. 1). Наскальные изображения людей-грибов также свидетельствуют о поклонении им народов, населявших Сибирь.
Рис. 1. Каменные статуэтки грибов
Однако сведений об истинной природе грибов, их биологии не было. Возникновение грибов после дождей связывали с ударами молнии. Появление грибов на листьях растений объясняли влиянием росы или продуктов выделения растений. В истории становления микологии как науки выделяют несколько этапов. Первый этап, продолжающийся до середины XIX в., связан с накоплением материала, описанием новых видов, попытками их классификации. Первые научные данные о грибах относятся ко второй половине XVI в. В этот период натуралист К. Клузиус (1526–1609),используя собственные сборы и материалы других исследователей, составил первую систематическую сводку о грибах. Бесценна его коллекция, состоящая из 221 акварельного рисунка грибов, известная под названием Кодекс Клузиуса (хранится в библиотеке Лейденского университета, в Голландии). Обычно грибы изучались попутно с другими организмами. Первым специалистом в области микологии считают итальянского ученого П. Микели. Совершенствуя оптические приборы, он сделал открытие (1729 г.), согласно которому грибы образуются в процессе прорастания мельчайших крупинок, названных позднее спорами. В результате грибы были причислены к царству растений. Так был развеян миф о загадочном возникновении шляпочных грибов. Знаменитый ученый К. Линней (1707–1778) внес свой вклад и в развитие микологии. Вначале он относил грибы к царству животных, обнаружив некоторое их сходство с полипами. Впоследствии он определил их в XXIV класс своей знаменитой системы, куда также были включены водоросли. Попытка Линнея систематизировать организмы способствовала возникновению новой науки о грибах – микологии. Большая работа по обобщению данных о грибах была проделана Х. Линком (1767–1850). Впоследствии, опираясь на накопленный материал по грибам, голландский исследователь Х. Г. Пирсон (1755–1836) и шведский ученый Э. М. Фриз (1794–1878) предприняли попытку систематизировать грибы. Эти ученые стали основоположниками систематики грибов, являясь представителями различных течений. Так, Пирсон стремился к формированию естественной группировки грибов, следуя воззрениям Ламарка. Фриз большое значение придавал анатомическим методам исследований, вслед за Линнеем придерживаясь создания искусственной системы. Э. Фриз предложил выделить грибы в самостоятельное царство. Эта идея в то время не нашла широкого распространения и была поддержана впоследствии лишь некоторыми учеными: Конардом (Conard, 1939), Б. М. Козо-Полянсим (1947) и др. Многие виды и роды, установленные этими учеными, сохранились до сих пор в микологической номенклатуре. В первой половине XIX в. активно ведутся исследования с целью выявления микобиоты различных регионов, параллельно изучаются филогения грибов, их строение, цитология. Большое значение приобретают исследования паразитических грибов. Этот период знаменуется деятельностью таких ученых, как А. И. Корда (1809–1849), Г. Л. Рабенсгорст (1806–1881), И. М. Барклей (1803–1889), М. К. Кук (1825–1914). Грибы России изучались сначала путешественниками. Даже у Линнея есть публикации (1737, 1792) о нахождении в России 155 видов грибов. Первые значительные микологические работы относятся к 1750 г. составлен список, включающий 430 видов грибов, собранных в окрестностях Петербурга. К 1836 г. Н. А. Вейнман (1782–1868) описал 1123 вида грибов России. Этого знаменитого ученого по заслугам считают первым русским микологом. Второй этап характеризуется подъемом в развитии микологии (со второй половины до конца XIX в.). Изучаются онтогенез и филогенез у грибов, исследуются циклы развития, особенно паразитических видов. В это время закладываются научные основы фитопатологии – науки о болезнях растений. Взоры микологов устремляются на исследование не только макроскопических, но и микроскопических грибов. Этот период связан с работами выдающихся учёных – братьев Тюлан во Франции, А. де Бари (1831–1888) в Германии. Так, Л. Тюлан (1815–1885) установил, что явление плеоморфизма характерно для всех групп грибов. Плеоморфизм – наличие различных последовательных спороношений в сложном жизненном цикле грибов. Это открытие является одним из самых крупных в микологии. До этого многие стадии спороношения одного и того же вида принимали за разные виды. А. де Бари является основоположником экспериментальной микологии и по праву считается отцом микологии. Он был автором первой филогенетической классификации грибов, основанной на признании их происхождения от водорослей. Ботанический институт в Страсбурге становится центром микологических исследований. Огромной заслугой А. де Бари явилось создание большой школы микологов и фитопатологов, среди которых было много русских ученых. Изучение видового разнообразия грибов в этот период не потеряло актуальности, исследования проводятся в различных уголках земного шара. Накопленный материал был обобщен П. Саккардо (1845–1920), который описал все известные к этому времени виды грибов земного шара. В 25 томах были представлены сведения о 74 323 видах. Большая роль в развитии микологии принадлежит О. Брефельду (1839–1925), разработавшему методы получения чистых культур грибов. В России Л. С. Ценковский (1822–1887) заложил основы изучения морфологии и циклов развития грибов и миксомицетов, его работы по этим вопросам считаются классическими. По отзывам современников, Л. С. Ценковский постепенно открывал перед наукой замечательный мир микроорганизмов. Он создал научные школы ботаников и бактериологов. Интересы М. С. Воронина (1838–1903), ученика де Бари, касались различных сторон микологии, его многочисленные работы связаны с изучением сложных явлений в жизни грибов. Он занимался исследованием капустной килы, ржавчины подсолнечника, биологии микоризных грибов. Появление большинства его работ вызвано практическими потребностями сельского хозяйства. М. С. Воронина по праву считают отцом русской микологии и основателем русской фитопатологии. Третий этап в развитии микологии характеризуется развитием физиологии и биохимии грибов (конец XIX – середина XX в.). На грибы обращают внимание не только микологи, но и физиологи растений, изучавшие у них различные физиологические процессы (дыхание, брожение, метаболизм). Многие исследования носят экологический характер, так как в ходе их выяснялось влияние факторов среды на онтогенез грибов (Г. Клебс и др.). С развитием техники появляется возможность изучения клетки грибов, её химического состава. С этой целью П. Данжаром, Р. Гернером, П. Клауссеном, применялся цитологический метод. Большое внимание уделяется исследованию биологических особенностей патогенных грибов, возбудителей болезней у растений, животных и человека. В России в этот период развиваются практически все известные направления микологии. Систематик и морфолог Н. В. Сорокин (1846–1909) известен своими работами в области изучения паразитических грибов растений и животных. Возникшие благодаря ему научные направления в области микробиологии развивались в опытах Ф. М. Каменского (1851–1912), впервые описавшего мицелий гриба на корнях растений (микориза); Ф. М. Породько (1877–1948), получившего за работу по изучению изменения бродильной активности дрожжей в зависимости от усиленного питания пептоном, степень кандидата наук по окончании университета; И. Л. Сербинова (1872–1925), автора учебника «Общая микробиология» (1916), который изучал бактериальные болезни растений, строение и биологию хитридиевых грибов, описал новые виды фитопатогенных бактерий и грибов. Выдающийся ученый А. А. Ячевский (1863–1932) исследовал видовое разнообразие грибов, а также ржавчинные и мучнисторосяные грибы, бактериальные и вирусные болезни растений. Основные его труды посвящены систематике и филогении грибов. Он является автором первого на русском языке определителя грибов (1897). Известна большая организаторская деятельность А. А. Ячевского. В 1902 г. в Петербурге им создана Центральная ботаническая станция, в 1907 г. – Бюро по микологии и фитопатологии при Министерстве сельского хозяйства, отдел микологии и фитопатологии (впоследствии лаборатория микологии им. А. А. Ячевского) при Институте опытной агрохимии. Под руководством А. А. Ячевского регулярно издавался сборник «Материалы по микологии и фитопатологии». Являясь профессором высших учебных заведений в Петербурге, он был известен своей активной просветительской деятельностью. В. А. Траншель (1868–1941) занимался в основном изучением биологии ржавчинных грибов, которые были собраны им лично или входили в многочисленные коллекции. Он предложил метод исследования разнохозяйности ржавчинных грибов, ныне используемый во всём мире. Л. И. Курсанов (1877–1954) исследовал морфологию и цитологию грибов, главным образом ржавчинных, взаимоотношения паразитных грибов и растений-хозяев. Им был внедрен цитологический метод в микологию. Его учебник «Микология» до настоящего времени популярен среди микологов. Н. А. Наумов (1888–1959), ученик А. А. Ячевского, продолжил исследование микобиоты различных регионов, в особенности Европейской части СССР, Средней Азии, Алтая, Дальнего Востока. Основные его работы посвящены систематике мукоровых грибов, им описано около 200 новых видов. Известны исследования учёного по фитопатологии, посвящённые явлениям и проблемам паразитизма, иммунитета, фузариоза и ржавчины хлебных злаков, килы капусты. Много времени Н. А. Наумов уделял педагогической работе как профессор Ленинградских высших учебных заведений. Им были написаны многие учебники и учебные пособия, переведенные на разные языки. Известнейший русский миколог А. С. Бондарцев (1877–1968) проводил микологические и фитопатологические исследования в различных районах СССР, опубликовал руководство «Грибные болезни культурных растений и меры борьбы с ними», долгое время бывшее единственным учебником по фитопатологии. Широко известен его капитальный труд «Трутовые грибы европейской части СССР и Кавказа». В ХХ в. микологические исследования во всех указанных направлениях осуществляются плеядой ученых и научными коллективами всех отделений Российской Академии наук и высших учебных заведений. Статьи по микологии публикуются главным образом в журналах «Микология и фитопатология» (с 1967 г.) и «Новости систематики низших растений» (с 1964 г.). Во второй половине ХХ в. благодаря работам Р. Уиттейкера (1969) и А. Л. Тахтаджяна (1970) грибы рассматриваются в ранге царства во всех современных системах. В этот период начинает складываться новый, четвертый, этап в развитии микологии, связанный с изучением генетики грибов. Именно потребности человеческого общества стимулировали развитие нового направления микологии: грибы, продуцирующие разнообразные биологически активные вещества – ферменты, антибиотики, фитогормоны, становятся популярны как объекты биотехнологии. Американские ученые, лауреаты Нобелевской премии Д. Бидл (1903–1989) и Э. Тейтем (1909–1975), открыв биохимические мутанты у сумчатого гриба Neurospora crassa, заложили основы биохимической генетики. Развитие этого направления шло от решения вопросов прикладного характера, связанных с селекцией грибов, используемых в биотехнологии, до выяснения вопросов теоретической микологии. В частности, поднимаются проблемы систематики грибов, филогении, изучения вида в онтогенезе и на популяционном уровне, его экологических особенностей. В последнее время становится популярной молекулярная систематика, или геносистематика, в основе которой лежит сравнение ДНК исследуемых организмов, что позволяет сопоставлять генотипы, а не фенотипы. На основе генного анализа в настоящее время пересмотру подвергаются все системы живых организмов, в том числе грибов.