Начинается с момента раскрытия цветка и опадения колпачка и опадения колпачка венчика и длится до завязывания ягод. Это важный этап в развитии генеративных органов. Во время цветения происходят следующие процессы: отрыв лепестков венчика от основания цветка, распрямление тычинок, сброс колпачка, растрескивание пыльников, высыпание пыльцы, падение её на рыльце пестика, набухание и прорастание пыльцы, двойное оплодотворение, образование зиготы и начало развития зародыша и семени, завязывание ягоды, опадение цветков и завязи. Все эти процессы для одного насаждения происходят в 8…14 дней, обычно в течение первых двух декад июня. Кусты, у которых почки распускаются раньше, раньше и цветут.
Для наступления фазы цветения требуется накопление суммы активных температур с начала вегетации в среднем около 380º С. Величина её специфична для каждого сорта, что необходимо учитывать при подборе сортов-опылителей. Фаза цветения коррелирует с фазой созревания ягод: чем раньше проходит цветение, тем раньше наступает созревание ягод.
Раннее цветение наблюдается при большей обеспеченности теплом района, участка произрастания, почвы и места в пределах одного насаждения. Цветение куста и соцветия наступает в разное время: раньше — у соцветий, расположенных ближе к поверхности почвы. Если на одном побеге располагается несколько соцветий, то раньше наступает цветение у соцветия, расположенного ближе к основанию побега, затем последовательно вверх. Раньше зацветают цветки, расположенные ближе к основанию соцветия. Из трёх бутонов, расположенных на последнем порядке ветвления соцветия, раньше распускается центральный, затем боковые. Длительность цветения одного соцветия составляет в среднем 4…9 дней, причём длительность цветения зависит от времени его начала: чем позже начинается цветение, тем интенсивнее и за более короткие сроки оно проходит. Указанные закономерности цветения проявляются в разновременности созревания ягод в пределах насаждения, куста и грозди, что необходимо учитывать при отборе средней пробы и определения сроков сбора урожая.
Бутоны раскрываются утром, между 6 и 11 ч, наиболее интенсивно — между 9…10 ч, затем процесс раскрытия бутонов затухает. Во второй половине дня между 15 и 16 ч наблюдается ещё одна волна раскрытия бутонов, но менее значительная. Связано это с изменениями температуры воздуха и его влажности в течение дня.
Вскоре после раскрытия цветка на рыльце пестика выделяется секреторная жидкость, к которой прилипает пыльца. Рыльце сохраняет способность к восприятию пыльцы в течение 4…6 дней после раскрытия цветка. Причём в эти дни наблюдается определённая суточная закономерность динамики: секреторная жидкость выделяется ритмично в ранние утренние и вечерние (17…18 ч) часы, пока не произойдёт опыления или не подсохнет рыльце цветка. После попадания пыльцы на рыльце оплодотворение происходит в течение суток, а затем рыльце пестика окрашивается в бурый цвет и образование секреторной жидкости прекращается. Начинается рост завязи. Некоторые авторы (К. Д. Стоев) отмечают наличие у цветков протерандрии.
В процессе цветения, опыления и завязывания ягод значительная часть цветков опадает в виде бутонов, распустившихся цветков и завязей. Это естественный процесс, и опадение до 40…80% от числа цветков, образовавшихся в соцветии, считается нормальным и приводит к образованию гроздей типичного сложения. Количество опавших цветков и завязей не имеет значения при условии сохранения в сформировавшейся грозди ягод соответствующего среднему показателю количества, характерного для данного сорта и обеспечивающих образование грозди типичного сложения и размера. Однако наблюдаются случаи и чрезмерного осыпания, обусловленные внутренними и внешними причинами.
Из внутренних причин наиболее важные — тип цветка, нарушения в строении андроцея и гинецея, нарушения гормонального баланса, недостаток питания, часто связанный с усиленным ростом побега, несущего соцветие и т. д. Влияние их можно ослабить или устранить проведением различных приёмов: предварительным (перед цветением) прищипыванием побега, обработкой регуляторами роста (ретарданты — ингибирование роста побега, гиббереллин — стимулирование роста завязи), подкормкой питательными элементами перед цветением и т. д. Правильное размещение побегов на шпалере в предыдущую фазу также создаёт благоприятные условия для оптимального перераспределения питательных веществ между элементами побега и одновременно способствует созданию лучшего фитоклимата в кроне куста. В случае образования нежизнеспособной пыльцы необходимо дополнительное опыление заранее собранной пыльцой.
Из внешних факторов важнейшее значение имеет температура, оптимальная величина которой для фазы цветения составляет 25…30º С. При температуре ниже 10…13º С пыльца не прорастает, а ниже 15…16º С цветение проходит вяло и прекращается рост тыльцевой трубки, что приводит к чрезмерному осыпанию, формированию большего числа ягод партенокарпических и стеноспермокарпических. Из-за прохладной, пасмурной, дождливой погоды в период цветения плохо раскрываются цветки, затрудняется перенос пыльцы и оплодотворение. Дожди смывают с рыльца секреторную жидкость и создают препятствия, позволяющие регулировать микроклимат, особенно температурный режим виноградника, — выбор более теплообеспеченных и защищённых от ветров мест, устройство защитных насаждений, правильное расположение рядов, открытость кроны куста, способствующая прямому попаданию солнечных лучей на соцветия и повышению их температуры. Чрезмерно высокая температура также может препятствовать опылению, особенно в сочетании с высокой сухостью воздуха и сильным ветром, что приводит к быстрому подсыханию рылец. Большое значение имеет достаточная влагообеспеченность, создающая условия для интенсивного питания, транспирации и фотосинтеза, однако нельзя поливать во время цветения, так как это снижает температуру.
В фазе цветения продолжается интенсивный рост побегов, листьев, усиков, пасынков, активно растут корни, работает камбий, начинается лигнификация стенок клеток побегов в нижней зоне. В пазухах листьев нижних узлов продолжается дифференциация зимующих глазков, для центральных почек которые наступает первый критический период в формировании генеративных органов.
Таким образом, фаза цветения отличается активным использованием для вегетативного и генеративного роста и развития виноградного растения питательных веществ, высокой интенсивностью процессов метаболизма. В эту фазу решается в значительной мере величина урожая текущего года, а также в определённой степени и будущего урожая.
Вторая фаза вегетации винограда наступает с начала распускания почек и длится до наступления цветения. С распусканием почек и появлением зелёных органов растение вступает в совершенно новое качество.
Распускание почек винограда наступает во второй половине апреля — начале мая в зависимости от условий года. Длительность фазы составляет 25…55 дней, начинается она с появления между распускающимися чешуйками верхушки побега. При это растягиваются зачаточные междоузлия, интенсивно делятся клетки в конусе нарастания и формируются новые узлы.
В период распускания почек и начала роста побегов продолжается дожифференциация зачаточных соцветий, формирование элементов цветка — андроцея, гинецея. В зависимости от обеспеченности питанием, гормонального статуса и некоторых других факторов достигается более полная реализация зачаточной плодоносности или наблюдается частичная редукция элементов соцветия.
В распускании почек и скорости роста побегов проявляются эволюционно обусловленные закономерности: строгая последовательность в развитии почек, которая выражается в том, что раньше начинают развиваться и распускаться более дифференцированные центральные почки, затем более дифференцированные замещающие и т. д., в последнюю очередь — спящие. На развитие почек большое влияние оказывает также пространственное расположение элементов куста и их частей, несущих почки. На вертикальных и наклонных стрелках преимущество в развитии имеют верхние почки, на горизонтальных стрелках почки развиваются равномерно, а при подвязке плодового побега в виде дуги, кольца преимущество получают почки, близкие к его основанию. Таким образом, изменяя пространственное расположение элементов куста, применяя различную длину обрезки и другие приёмы, можно управлять процессом распускания почек.
Раньше распускаются почки на коротко обрезанных плодовых побегах, чем на длинно обрезанных. Более раннее развитие отмечено также у почек, расположенных в более тёплых микроусловиях, в частности, в теплице раньше распускаются почки, расположенные близко к отопительным трубам.
На сильных кустах развивается больше точек роста. Однако количество образовавшихся побегов, как правило, больше, чем число полноценных побегов, которое может развить куст. Поэтому число побегов необходимо регулировать при помощи обломки.
На распускание почек и начало роста побегов влияют внешние факторы: температура, влажность воздуха и почвы, поступление воздуха во внутренние части глазка. С повышением температуры интенсивность распускания почек нарастает, стимулирует распускание почек достаточное водоснабжение, высокая влажность воздуха, хорошая аэрация.
После распускания почек рост побегов сперва идёт медленно, затем нарастает и достигает максимума в период, близкий к цветению. К концу фазы побеги достигают до 60% своей конечной длины. Одновременно с ростом стебля побега начинается и рост других органов, расположенных на нём, — соцветий, листьев, усиков, пасынков, почек.
Существует корреляция между ростом побега и ростом органов, расположенных на нём. С появлением зелёных органов побега начинается интенсивная транспирация и прекращается «плач».
В период распускания почек, а затем в начале роста побегов затрачивается много питательных веществ, так как фотосинтез ещё отсутствует или слабый. Для этого мобилизуются запасные питательные вещества из корней, стебля. По мере роста побега интенсивность фотосинтеза нарастает и всё большую долю от общих затрат на построение стебля составляют продукты ассимиляции текущего года.
Одновременно с удлинением побегов и корней, ростом других органов постепенно усиливается деятельность камбия — происходит рост стебля и корня в толщину. Один из важнейших процессов этой фазы — создание мощного ассимиляцонного аппарата. По некоторым данным, к концу фазы на побегах образуются до 80% общего количества листьев. Все эти процессы требуют больших затрат питательных веществ, поэтому большое значение, особенно до начала продуктивной работы листьев, имеют запасы, накопленные в корнях и стебле. С началом интенсивной деятельности листовой поверхности (фотосинтез, транспирация) резко усиливается сосущая сила и рост корней, интенсивность поглощения воды и питательных веществ корнями.
Оптимальная температура для роста побегов 25…30º С. Более высокие температуры для активного роста требуют высокой влажности почвы (80…85% НВ) и особенно воздуха. При недостаточной влажности почвы побеги резко замедляют рост, верхушки их выпрямляются. Температуры ниже 12…10º С приводят к прекращению роста. Удлинение светового дня стимулирует рост побегов. Тормозят рост побегов ветры: удаляют с виноградника влажный воздух, усиливают транспирацию, снижают температуру листьев, препятствуют лучшему использованию солнечных лучей, изменяя положение листьев, и в результате ослабляют рост побегов.
В развитии виноградного растения с точки зрения формирования продуктивности и выполнения агротехнических приёмов можно выделить наиболее важные этапы.
При появлении первого симподиального узла устанавливается плодоносность побегов, что даёт возможность провести обломку лишних и бесплодных побегов и установить оптимальную нагрузку.
Проведение агробиологических учётов, которые позволяют рассчитать величину возможного урожая. При этом в случае перегрузки часть соцветий, начиная с недоразвитых, удаляются.
По достижении растущими побегами при их вертикальном ведении 35…50 см проводят их первую подвязку к шпалере.
При большой нагрузке кустов основными побегами прищипывают жирующие на раннем этапе роста для получения пасынков с параметрами нормальных побегов.
За 1…2 недели до прогнозируемого начала цветения прищипывают плодоносные побеги у сортов, склонных к осыпанию бутонов, цветков и завязей, для увеличения массы грозди. С этой же целью можно применять опрыскивание растворами регуляторов роста ретардантного действия.
По мере нарастания вегетативной массы куста необходимо проводить профилактические мероприятия по защите виноградников от болезней и вредителей.
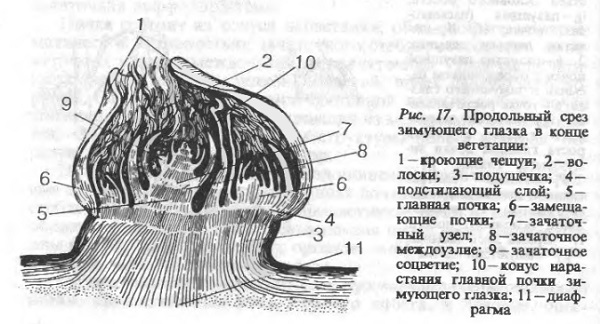
В процессе роста в пазухах листьев побега закладываются и начинают развиваться пазушные почки. В этот период происходит начало развития наиболее важных, с точки зрения формирования урожая будущего года, глазков нижней зоны побега, которые остаются после обрезки перед следующей вегетацией. В конце этой фазы почки почки подходят близко к началу первого критического периода в своём развитии или он для них уже наступает. Выполнение указанных и других агротехнических мероприятий способствует созданию условий для оптимизации процесса морфогенеза почек и закладки зачаточных соцветий.
Сокодвижение — первая фаза вегетационного периода, длится с начала весеннего сокодвижения до начала распускания почек. Внешне эта фаза проявляется в выделении на поверхности срезов или поранений стебля пасоки. Пасока представляет собой прозрачную жидкость с плотностью при температуре 15 С, равной 1,0007. Содержание сухого вещества в одном миллилитре — 1…2 мг, две трети которого органические вещества (сахара, азотистые вещества) и одна треть минеральные (К, Са, Р, Mg и др.). Реакция — слабокислая, рН 6,8. Установлено также: в более поздний период «плача» концентрация пасоки, особенно органических веществ, повышается.
Появление пасоки свидетельствует об окончании периода покоя и начале активной жизнедеятельности куста. Это связано с повышением температуры почвы и возобновлением процессов активной деятельности корневой системы. Перед началом вегетации почки и камбий находятся в состоянии вынужденного покоя, а у корней вообще отсутствует фаза глубокого покоя. Уровень температуры почвы, необходимый для начала сокодвижения, у сортов различного происхождения разный. У сортов вида Витис винифера сокодвижение начинается при температуре почвы 7…9º С, а у вида Витис амурензис и некоторых американских видов при более низкой температуре — 6…8º С. При достижении такой температуры усиливается дыхание в корнях, высвобождается энергия для их роста, активно поглощается вода из почвы и подаётся по сосудам древесины в виде пасоки в надземную часть под давлением 1,5 атм. С повышением температуры почвы интенсивность сокодвижения и «плача» возрастает, особенно при температурах выше 12º С. При наблюдении за динамикой весеннего сокодвижения винограда установлено, что интенсивность «плача» медленно нарастает, достигает максимума, а затем быстро падает. Главенствующую роль в прохождении фазы сокодвижения и интенсивности «плача» играет температура почвы. В фазу сокодвижения «плач» продолжается даже тогда, когда температура воздуха снижается ниже 0º С. Однако, если температура почвы снижается ниже указанных выше уровней начало сокодвижения, «плач» прекращается.
Начало и интенсивность «плача», а также величина выделенной пасоки зависят и от других факторов. При достаточном количестве влаги в почве «плач» интенсивный, при недостатке — слабый, а при критической нехватке влаги «плач» может и вовсе отсутствовать. Отсутствие его может быть также следствием подмерзания корневой системы. Более сильные кусты с мощной корневой системой, охватывающей больший объём почвы, отличаются интенсивным «плачем», слабые кусты — незначительным. Наряду с отмеченными выше видовыми особенностями существуют также и сортовые особенности «плача». На кустах быстропрогреваемых участков и почв, а также участков, где основная масса питающих корней располагается близко к поверхности, «плач» начинается раньше.
Выделение пасоки на поверхности ран зависит от времени их нанесения. Более обильный и ранний «плач» отмечается на более поздних ранах, особенно при обрезке после начала сокодвижения. Потеря пасоки ослабляет кусты, поэтому обрезка должна быть закончена до начала сокодвижения. Особенно ослабляются кусты при значительных потерях пасоки в более поздний период «плача», так как содержание органических веществ в этот период выше, чем в начальные этапы «плача».
Выделение пасоки на поверхность раны постепенно прекращается вследствие закупорки сосудов камедистыми веществами, которые образуются клетками древесинной паренхимы и различными микроорганизмами. При ежедневном обнавлении срезов на 1…2 см можно добиться продолжения выделения пасоки и установить определённые закономерности, касающиеся её количества. Так, по данным А. С. Мержаниана, количество выделяемое одним кустом пасоки составляло при обновлении срезов в различных условиях 1,4…5 л, а без обновления — 0,2…2,0 л. В отдельных случаях очень сильные кусты при обновлении срезов выделяли до 16 л пасоки.
Наряду с внешними проявлениями начала вегетации в виноградном растении происходят сложные процессы метаболизма. В корнях и многолетних частях кустов происходит распад сложных органических соединений, которые передвигаются по восходящему току к зонам меристематической активности. Активизируются ферменты, стимулирующие процессы метаболизма, а также деятельность камбия и других меритем. В результате увеличиваются в размерах клетки зачаточных междоузлий почек зимующих глазков, что приводит к набуханию глазков, раскрытию чешуй и появлению элементов побега, заложенных в предыдущую вегетацию. Этот момент считается временем окончания первой фазы вегетации.
Длительность фазы «плача» зависит от сорта, температуры и влажности почвы и составляет 10…50 дней. Для завершения фазы сокодвижения и начала второй фазы необходима сумма активных температур в 145…258º С, что является результатом приспособления и предотвращает распускание почек до наступления устойчивых положительных температур. Следует отметить, что для винограда вида Витис амурензис и некоторых американских видов. Быстрое прохождение фазы сокодвижения может привести в районах, где наблюдаются весенние заморозки, к повреждению кустов.
Завершающий этап фазы сокодвижения — один из важнейших периодов в формировании генеративных органов. В этот период наряду с началом развития почек возобновляется додифференциация соцветий. При неблагоприятных условиях может происходить редукция элементов соцветия с образованием переходных к усику форм, а иногда полное превращение их в усик.
В целом фаза сокодвижения является одной из важнейших в годовом цикле виноградного растения. В начале её происходит глубокая физиологическая и биохимическая перестройка организма, в процессе которой кардинально меняется гормональный статус виноградного растения при переходе от состояния покоя к вегетации. Отсюда важность комплекса мероприятий по уходу за кустами. Некоторые из важнейших агроприёмов необходимо проводить в эту фазу. Это ремонт шпалеры, сухая подвязка (особенно плодовых побегов), искореняющее опрыскивание против зачатков вредителей и болезней, прививки на месте, основное внесение азота, глубокое рыхление и некоторые другие.
В процессе онтогенеза виноградное растение ежегодно проходит годичный (малый) цикл — совокупность морфологических, анатомических, физиологических и биохимических изменений и процессов, связанных с сезонными изменениями в течение года. При развитии виноградного растения в малом цикле, наряду с внешними изменениями в строении растения, происходят сложнейшие процессы перестройки анатомии, физиологии, биохимии, синтеза сложных соединений и расщепления, усвоения и мобилизации элементов среды обитания, вступления в покой и выхода из него, старения и обнавления и т. д. Ряд процессов строго следует друг за другом, другие протекают параллельно, сливаются. Процессы развития в малом цикле, так же как и в большом жизненном цикле, генетически обусловлены. Направление и интенсивность их подчинены воздействию факторов среды. Ритмы процессов метаболизма, развития в годовом цикле удивительно приспособлены и соответствуют динамике основных факторов среды, главным образом света и температуры, в течение года в районах возникновения и эволюции виноградного растения. Вместе с тем растение винограда возделывается далеко за пределами экологического оптимума, а габитус и ценоз культурного растения совершенно не соответствуют таковым естественно произрастающего растения. Знание процессов развития растения винограда в годовом цикле, оптимального уровня факторов для каждой фазы позволяет управлять формированием устойчивости, продуктивности растений и качества продукции.
В полосе субтропического и умеренного климата в годовом цикле винограда наблюдается два периода: вегетационный период и период покоя.
Вегетационным называется период, в течение которого проявляются активные процессы жизнедеятельности куста, — рост, развитие, плодоношение и т. д. Это период начинается весной с начала сокодвижения («плач»), затем происходит распускание почек, рост побегов, вызревание побегов, созревание урожая, прекращение роста, листопад. В процессе прохождения вегетационного периода выделяют фазы, на каждой из которых растение находится в качественно новом состоянии, требующем специфического уровня и набора факторов внешней среды. Существует несколько подходов к разделению вегетационного периода на фазы.
Некоторые авторы выделяют три фазы — роста, плодоношения и вызревания побегов, другие — четыре, увязывая их с биохимическими процессами. В нашей стране принято разделять вегетационный период на шесть фаз: сокодвижение, рост побегов и соцветий, цветение, рост ягод, созревание ягод, вызревание побегов и листопад. Такой вариант оптимален, так как выделены наиболее важные внешние проявления каждого этапа развития растения в процессе вегетации. Это разделение позволяет планировать различные агротехнические мероприятия в соответствии со спецификой состояния растения на каждом этапе вегетации. По данным фенологических наблюдений за несколько лет устанавливают средние календарные сроки наступления и продолжительности фаз, подготавливают и проводят агротехнические мероприятия. Сроки наступления отдельных фаз, продолжительность их зависят от почвенно-климатических условий, сорта, топографических условий местности, возраста кустов, особенностей погодных условий в данную вегетацию.
Каждый современный представитель растительного мира, в том числе и виноград, является продуктом эволюционного развития. В результате естественного и искусственного отбора достигли современного уровня эволюции и сохранились те формы, которые наиболее соответствуют условиям внешней среды и хозяйственным нуждам человека. В генетическом аппарате растений выработана и закреплена информация, которая является основой для индивидуального развития каждой особи. В процессе онтогенеза — индивидуального развития виноградного растения от его зарождения до гибели — эта наследственная информация реализуется. Наследственная основа каждой особи определяет только общий порядок и направление морфогенетических процессов, а реализуются они под действием внешней среды. Специфика индивидуального развития определяется генетически обусловленной нормой реакции особи. Существенное влияние на процессы онтогенеза культурных растений, в том числе и винограда, оказывает уход за ними, т. е. антропогенные факторы.
Виноградное растение обладает высокой способностью онтогенетической адаптации к условиям внешней среды, экологической пластичностью. Но вместе с тем наиболее плотная реализация генетического потенциала продуктивности, физиологической продолжительности жизни осуществляется в условиях, максимально соответствующих потребностям растения, выработанным в процессе филогенеза.
В процессе онтогенеза растение проходит большой жизненный цикл — совокупность изменений в морфологии и функциях в течение жизни — и ежегодно малый (годичный) цикл. Онтогенез семенного растения начинается с момента образования зиготы в результате оплодотворения яйцеклетки на материнском растении, а особи, полученной вегетативно, — с момента закладки почки на побеге, используемом для размножения. Виноградное растение является поликарпическим и характеризуется довольно длительным онтогенезом. Установлено, что в благоприятных условиях условиях произрастания, при одиночном стоянии в диком или культурном виде виноградное растение может достигать 300-летнего возраста. В культуре продолжительность жизни виноградного растения значительно короче и в зависимости от экологических условий выращивания, способа культуры, ухода составляет 20…60 лет. В Западной Европе средняя продолжительность жизни виноградников составляет около 30 лет, а в нашей стране меньше, что связано с более суровым климатом. При укрывной культуре продолжительность жизни насаждения ещё меньше.
На протяжении онтогенеза изменяется морфология, структура виноградного растения, направленность и интенсивность физиологических и биохимических процессов. В специфике прохождения этих процессов в течение онтогенеза устанавливается этапность, связанная с возрастом особи. Существуют различные взгляды на дифференциацию онтогенеза и определение этапов. Для виноградного растения принято разделение жизненного цикла на 4 этапа: эмбриональный, ювенильный, продуктивный и старения. На каждом из этих этапов виноградному растению присуща определённая специфика его состояния.
Эмбриональный этап у растений, полученных из семян, длится с момента образования зиготы до прорастания семени и образования первого настоящего листа. У вегетативно размноженных — с момента закладки почки до её прорастания. Важной спецификой этого этапа является то, что значительная часть его протекает на материнском растении. Следовательно, условия существования материнского растения определяют жизнеспособность будущих дочерних растений, интенсивность их развития в начальной стадии онтогенеза. Поэтому на маточниках винограда необходим специфический уход.
Особенностью развития на эмбриональном этапе является то, что и семена и почки после формирования вступают в состояние покоя, связанного с их гормональным балансом. Для преодоления покоя семян имитируют условия, в которые попадают семена в естественных условиях: содержат длительное время во влажном виде при низких положительных температурах. Это снимает физиологические барьеры при прорастании семян. В последующем при благоприятных условиях активируется дыхание, что высвобождает энергию, что необходимую для прорастания, нерастворимые вещества эндосперма превращают в растворимые и вовлекаются в метаболизм. Прорастающий зародыш потребляет эти вещества. Сначала образуется стержневой корень, затем семядольные листья выносятся на поверхность субстрата, в результате деятельности конуса нарастания почек образуются листья. При появлении первого настоящего листочка заканчивается эмбриональный этап сеянца.
Почки, из которых формируется надземная часть будущего дочернего растения, в процессе своего развития проходят несколько этапов покоя до полной дифференциации: предварительный покой, с августа — глубокий, а с ноября — вынужденный. При использовании для размножения почек (зелёные черенки) на этапе предварительного и вынужденного покоя происходит быстрое распускание почек, что иногда приходится задерживать (кильчевание).
Длительность эмбрионального этапа составляет у семенных растений от 6 месяцев, а вегетативно размноженных — от 3 месяцев.
Ювенильный этап длится с момента образования первого листа до вступления растения в плодоношение. На этом этапе происходит постепенное нарастание объёма корневой системы, вегетативной массы куста, что обеспечивает переход в качественно новое состояние — плодоношение. Семенные растения отличаются более длинным ювенильным этапом — первые генеративные органы у них образуются на 3…7-й год в зависимости от происхождения (сеянцы сортов восточной группы более поздно вступают в плодоношение), условий выращивания. При помощи различных агротехнических мероприятий можно победить сеянцы к более раннему плодоношению. К таким приёмам относится прививка сеянца в семядольном состоянии на взрослый куст, круглогодичная вегетация в теплицах, усиленное питание и др. В целом всё то, что усиливает растение, стимулирует образование вегетативной массы, побуждает к закладке генеративных органов. Этими приёмами пользуются для ускорения селекционного процесса, быстрой оценки гибридного потомства.
Полученные вегетативным путём растения способны очень быстро (2…4 года) формировать генеративные органы. Однако первые годы насаждения должны быть посвящены быстрому созданию выбранной формы. В этот период усилия направлены на создание вегетативной массы куста, формированию штамба, рукавов, рожков, плодовых побегов ( скелета куста). Обычно на это уходит 3…6 лет жизни насаждения в зависимости от природно-климатических условий, ухода, выбранной формы, биологических особенностей сортов. В некоторых случаях для получения полноценных побегов, предназначенных для создания элементов куста, практикуется удаление сформировавшихся генеративных органов. Важнейшим принципом в период формирования куста должно быть постепенное наращивание нагрузки кустов глазками и побегами по мере увеличения вегетативной массы. На этом этапе наряду с ростом у вегетативно размноженных растений происходит и частичное плодоношение. Следует отметить, что молодые растения на этапе роста имеют более длительный период вегетации, у них затягиваются ростовые процессы, наблюдается более позднее вступление в состояние покоя, что приводит к формированию более низкого уровня зимостойкости. Молодые растения более чувствительны к дефициту влаги, питания, к вредителям и болезням. Поэтому необходимо проводить мероприятия по своевременной и качественной подготовке к перезимовке, обеспечению оптимального водно-питательного режима, эффективной защите от вредителей и болезней.
Продуктивный этап для виноградных растений наступает с окончания формирования кустов и вступления в полное плодоношение. Этот этап самый длительный: его продолжительность составляет десятки лет. Надземная часть виноградного куста полностью сформирована и осваивает всё воздушное пространство, отведённое ему согласно схеме посадки, однако корневая система продолжает осваивать площадь питания. На этом этапе рост сочетается с ежегодным плодоношением. Оба эти процесса сосредоточены на одном органе — плодоносном побеге. Для обеспечения высокой продуктивности кустов, предотвращения периодичности плодоношения необходимо создавать условия для гармоничного сочетания роста и плодоношения. Для этого используют различные агротехнические мероприятия: удобрение, орошение, установления оптимальной нагрузки кустов побегами и урожаем, создание оптимального фитоклимата, предотвращение развития болезней и вредителей, регулирование роста и плодоношения путём проведения операций с зелёными частями куста, применения регуляторов роста и др.
Обеспечение гармоничного сочетания роста и плодоношения создаёт предпосылки для максимального продления продуктивного этапа. Молодые кусты до 7…8 лет отличаются сильным ростом, затем сила роста побегов постепенно ослабевает. Высокий уровень продуктивности кустов сохраняется ряд лет, затем постепенно снижается. На длительность этапа плодоношения влияют экологические условия, сортовые особенности, агротехника. В благоприятных климатических и почвенных условиях кусты винограда имеют длительный продуктивный этап, а при неблагоприятных — быстро стареют и теряют продуктивность.
Этап старения растения обусловлен продолжительностью функционирования проводящих сосудов и нарушениями в обмене белков и других веществ. Происходит закупоривание проводящих сосудов тиллами, сначала затрудняющее нормальное функционирование сосудисто-проводящей системы. Затем наступает полное закупоривание сосудов. В результате нарушается механизм транспорта воды и питательных веществ, что приводит к отмиранию тканей. Эти ткани пропитываются тёмно-окрашенными веществами и формируют ядровую древесину. На многолетних элементах куста ядровая древесина охватывает центральные более ранние годичные кольца, затем этот процесс перемещается к периферии. В более молодом, активном состоянии выключение из системы корнелистовой связи более старых частей проводящей системы компенсируется образованием молодых элементов проводящей системы сосудов благодаря деятельности камбия.
Нарушение корнелистовой связи при закупорке сосудов ослабляет рост, уменьшает ассимиляционную поверхность, что снижает количество вырабатываемых продуктов фотосинтеза и их поступление в многолетние части и корни. Это приводит к ослаблению роста корней и уменьшению поглощения из почвы и т. д.
Существуют определённые закономерности в старении кустов и органов. Чем сильнее рост кустов, тем позже они достигают максимальной продуктивности и имеют большую долговечность. Слаборастущие кусты раньше достигают максимальной продуктивности, быстрее стареют и раньше заканчивают жизненный цикл. Кусты средней силы роста занимают промежуточное положение.
Органы скелетной части куста имеют разную долговечность. Наибольшая долговечность характерна для подземного штамба, затем для надземного штамба, рукавов, рожков. Инактивация сосудов тиллами в зоне укрывной культуры начинается в рукавах в возрасте 2…3 лет, подземном штамбе — 10…15 лет, скелетных корнях — старше 18…20 лет. Ядровая древесина охватывает годичные кольца, расположенные, считая от периферии у рукавов, после 5…7-го годичного кольца, у подземного штпмба — после 18…25-го, у скелетных корней после 35…39-го. Установлено, что в указанных условиях сосуды функционируют в рукавах не более 4…5 лет после образования годичного кольца. Образование ядровой древесины резко активизируется при нанесении ран, заболеваний пятнистым некрозом и других повреждениях. Следует также отметить, что с возрастом ослабляется и деятельность камбия, что приводит к формированию меньшего количества проводящих элементов, более узких годичных колец, т. е. к необратимым процессам старения. Восстановлению продуктивности в этих условиях помогают приёмы омоложения рукавов и других элементов куста. Для их замены используются побеги, образовавшиеся из спящих почек. Иногда проводят омоложение на «чёрную головку» удаляя всю надземную часть куста и формируя новые элементы куста из порослевых побегов. При снижении продуктивности кустов ниже экономически обоснованных пределов насаждения винограда раскорчёвывают.
Важнейшим критерием характеристики отдельных сортов, агроприёмов, кустов и насаждений является продуктивность — способность формировать определённый биологический (биологическая продуктивность) и хозяйственный (хозяйственная продуктивность) урожай.
Хозяйственную продуктивность (урожай) составляет масса урожая гроздей (на маточниках — однолетних черенков) с единицы площади насаждения или куста. Хозяйственная продуктивность виноградника (урожайность) складывается из суммарной хозяйственной продуктивности составляющих его кустов и может быть потенциальное, эмбриональной и фактической.
Потенциальным называется урожай, который может быть получен с растения в идеальных условиях на основе 100%-й плодоносности зимующих глазков. Потенциальная продуктивность зависит от конструкции насаждения, которая определяет коэффициент полезного действия фотосинтетической активности радиации. КПД ФАР обычных промышленных виноградников со шпалерно-рядовой посадкой составляет около 1,5%, что соответствует 35…40 т/га урожая гроздей.
Эмбриональный урожай рассчитывается на основе изучения эмбриональной плодоносности (числа зачаточных соцветий на зимующий глазок) и их развитости в период покоя.
Фактический урожай кустов и урожайность виноградников, как правило, ниже эмбриональных и тем более потенциальных уровней. В процессе формирования фактической урожайности шпалерно-рядовых виноградников КПД ФАР составляет 0,3…1,0%, что связано недостаточным уровнем отдельных факторов. Создание условий для максимальной реализации эмбриональной урожайности и приближения уровней фактической и эмбриональной плодоносности к потенциальной является важнейшей проблемой современного виноградарства. Урожай куста и урожайность виноградника складываются из ряда структурных элементов — показателей продуктивности, на основе анализа которых создаются возможности направленного регулирования продуктивности и качества.
К ним относится число побегов на кусте и на единицу площади, процент плодоносных побегов, коэффициенты плодоношения и плодоносности, масса грозди. В связи с тем, что урожай винограда формируется только на побегах, развивающихся на кустах в текущем году, количество их на кусте и на единице площади будет определять продуктивность куста и насаждения.
Число побегов на га изменяется от тыс. до 200 тыс. шт. в зависимости от природно-климатических особенностей культуры, сорта, формы, агротехники, возраста насаждений. Максимальное приближение фактической урожайности к потенциальной обеспечивается при оставлении 25…30 побегов на погонный метр ряда при их вертикальном размещении и 40…50 — при свободном свисании (А. Г. Амирджанов).
Величина другого показателя продуктивности — процента плодоносных побегов — зависит от биологических особенностей сорта, условий культуры, ухода.
Для сортов восточной группы характерно относительно более низкое значение этого показателя по сравнению с сортами западно-европейской группы и группы бассейна Чёрного моря.
Побеги, образовавшиеся из центральных почек зимующих глазков, имеют более высокий процент плодоносных побегов по сравнению с побегами, образовавшимися из замещающих почек; плодоносные побеги из спящих почек развиваются очень редко.
Более высокий процент плодоносных глазков характерен для побегов, развившихся из центральных, а не замещающих почек. Зная закономерности формирования зимующих глазков, закладки и дифференциации в них генеративных органов, можно направленно влиять на процент плодоносных побегов путём применения различных агротехнических приёмов (удобрение, орошение, система формирования, операции с зелёными частями куста и др.). Особо важное значение имеет создание благоприятных условий в критические моменты развития генеративных органов — образование первого симподиального узла на зачаточном побеге центральной почки (период, близкий к фазе цветения), весенняя додифференциация соцветий (период распускания почек). Положительно влияют на плодоносность почек и побегов, развивающихся из них, подкормки, прищипывание, чеканка. Улучшение освещённости в кроне куста способствует повышению плодоносности почек и побегов. Установление на кусте оптимальной нагрузки также стимулирует плодоносность почек и побегов по сравнению с перегрузкой и недогрузкой кустов. Существенно влияет на процент плодоносных побегов состояние растения в период распускания почек. Более высокий процент плодоносных побегов наблюдается при медленном распускании почек. Быстрое распускание приводит к негативному развитию корневой системы и почек, вследствие чего часть зачаточных соцветий или их отдельные части редуцируются в усики. Задержка распускания путём применения регуляторов роста повышает процент плодоносных почек (З. Я. Молчанова). Сухая подвязка в более поздние сроки приводит к снижению процента плодоносных побегов и формированию большей доли соцветий неполного развития. Этот агроприём следует проводить в начале сокодвижения.
Важным показателем продуктивности является число соцветий (гроздей) на один плодоносный побег — коэффициент плодоносности. На одном плодоносном побеге обычно развивается 1…3 соцветия. Этот показатель является важным сортовым признаком. У крупногроздных сортов восточной группы на одном плодоносном побеге развивается 1, реже 2 соцветия, а у сортов других групп 2…3. На величину коэффециента плодоносности влияют условия в период закладки и развития зачаточных соцветий. Определяющими являются условия, при которых развиваются нижние 2…3 симподиальных узла эмбрионального побега центральной почки зимующего глазка. Влияет на коэффициент плодоносности и нагрузка куста урожаем. Улучшая питание формирующихся глазков путём прищипывания побегов, чеканки, повышения уровня водно-питательного режима, изменяя пространственную ориентацию побега, можно повысить коэффициент плодоносности побегов. Неблагоприятные условия перезимовки, гибель части центральных почек приводит к увеличению в структуре прироста доли побегов из замещающих и спящих почек и снижению коэффициента плодоносности. Существенно влияют и условия, при которых генаративные органы проходят второй критический момент — распускание почек весной следующего года.
Коэффициент плодоношения рассчитывается как отношение числа соцветий на кусте к общему числу развившихся побегов. Он изменяется в зависимости от сорта от 0,2 до 2, иногда и выше. На его величину влияют погодные условия, почва, применяемая агротехника и т. д.
Важным показателем, характеризующим продуктивность виноградного растения является средняя масса грозди. Для разных сортов он относительно постоянен. Средняя масса колеблется от 50…100 г до 1 кг и более. Более крупные грозди характерны для сортов восточной группы. Влияют на величину массы грозди условия произрастания и применения агротехники: нагрузка кустов урожаем, уровень питания, применение регуляторов роста. Особо важное значение имеет режим влажности: при более высоком уровне влагообеспеченности формируются более крупные грозди. Средняя масса грозди — один из ключевых элементов при прогнозировании и программировании урожаев винограда. При этом необходимо в условиях данной местности, участка, у данного конкретного сорта в течение нескольких лет определять массу грозди на основании взвешивания большого количества гроздей без выбора.
В последние годы для характеристики продуктивности сортов и агротехнических приёмов используют показатель продуктивности побега (А. Г. Амирджанов). Продуктивность побега — это масса урожая гроздей в расчёте на один развившийся на кусте побег. Рассчитывается путём умножения средней массы грозди на коэффициент плодоношения (Пп=К x Г). Средняя величина продуктивности побега для сорта, рассчитанная за ряд последних лет, называется индексом продуктивности сорта и служит важным ампелографическим признаком.
После цветения, опыления, оплодотворения, завязывания и осыпания излишних цветков и завязей соцветие винограда превращается в соплодие, которое носит название гроздь. Гроздь состоит из ножки грозди (гребненожка), гребня и ягод.
Ножка грозди формируется из ножки соцветия после завязывания ягод и представляет собой центральную ось от места прикрепления к побегу, несущему гроздь, до места отхождения первого ответвления. Для разных сортов винограда характерна различная длина гребненожки, что имеет значение для съёма урожая. В зависимости от сорта в процессе развития грозди ножка остаётся травинистой или одревесневает.
Гребень является скелетной частью грозди и состоит из главной оси, боковых ответвлений разных порядков (обычно до 2…3, реже до 4-х порядков), которые заканчиваются плодоножками. У некоторых сортов гребень остаётся при созревании ягод травянистым, ломким, у других одревесневает. Доля гребня в общей массе грозди составляет 1…8,5% (в среднем 3,5%) и изменяется в зависимости от сорта, условий формирования грозди, степени зрелости ягод, а доля в объёме грозди — до 30%. Прочность элементов гребня, их устойчивость к усыханию и повреждению болезнями, запасы питательных веществ в них, сила прикрепления ягод к плодоножкам определяют способность гроздей к хранению и транспортировке. Химический состав гроздей влияет на качество вина.
Все вышеуказанные признаки гребня характерны для определённых сортов и изменяются в зависимости от условий внешней среды. Обычно при производстве вина, сразу после поступления урожая на переработку ягоды отделяют от гребней, чтобы получить более мягкие, тонкие вина. Особенно ухудшает качество вина брожение в присутствии травянистых невызревших гребней — вина образуются жидкие, с травянистым, гребневым привкусом. При производстве вина по кахетинскому способу брожение осуществяют в присутствии гребней, но они должны быть хорошо вызревшими. Вина при этом получаются с большим содержанием дубильных веществ, терпкие, полные, с большей Р-витаминной активностью. В некоторых случаях получают путём пресования гребневое сусло, его сбраживают и используют для получения спирта или мадеры. Гребни после отделения от ягод идут на органическое удобрение.
Гребень заканчивается плодоножкой, которая формируется из цветоножки. Плодоножка в верхней части расширяется в подушечку, к которой крепится ягода. Через плодоножку в ягоду проникают сосудистые пучки, которые образуют при отрыве от ягоды кисточку, имеющую характерный для сорта тип (размеры, форму, окраску). Степень развития сосудистых пучков определяет прочность прикрепления ягоды к плодоножке. На поверхности плодоножек и подушечек образуются бородавки в виде выростов.
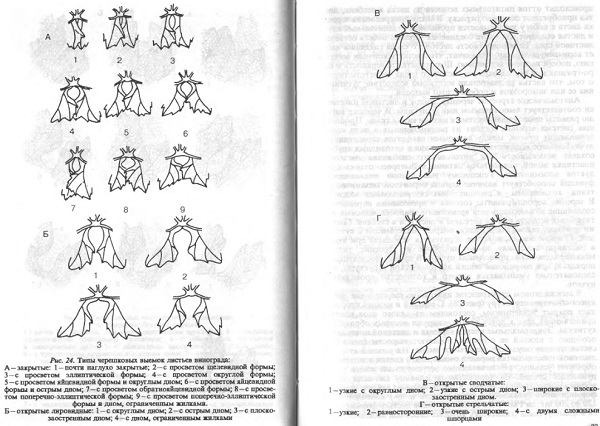
Характер расположения элементов ветвления гребня, их развитость, а также количество и размеры ягод определяют такие морфологические признаки гроздей, как размеры, форма и плотность. Эти показатели определяются сортом, однако на них влияют и условия формирования гроздей. Величина грозди характеризуется её длиной или массой. Очень крупными считаются грозди длиной более 26 см, крупными — 18…26, средними — 10…18 и менее 10 см — мелкими. Крупные и очень крупные грозди, имеющие ширину в 2 и более раза меньшую, чем длина, широкими — имеющие ширину, примерно равную 2/3 длины и очень широкими, если ширина приблизительно равна длине.
Каждому сорту соответствует своя специфическая форма грозди. Всё существующее многообразие форм гроздей в пять типов форм: цилиндрическая, коническая, цилиндро-коническая, крылатая и ветвистая. У крылатого типа сильно развито первое ответвление грозди, которе превращается в «крыло», что характерно для некоторых сортов.
Важным морфологическим показателем грозди является плотность. Различают очень плотные (ягоды так плотно располагаются, что изменяют свою форму), плотные (ягоды располагаются плотно, без просветов, но не изменяют форму друг друга), средней плотности (между ягодами имеются промежутки, но грозди не изменяют форму на ровной твёрдой поверхности при изменении положения), рыхлые (изменяют форму на ровной твёрдой поверхности при изменении положения), очень рыхлые (видны ответвления грозди, основные ответвления свободно располагаются в одной плоскости). Плотность грозди определяется длиной отдельных разветвлений, длиной плодоножек, количеством завязавшихся ягод, размерами отдельных ягод. Для столовых сортов предпочтительны грозди средней плотности, так как они более транспортабельны и лёжки.
Для технологической характеристики используют и другие показатели: масса грозди, число ягод в грозди, показатель строения грозди (отношение массы ягод в грозди к массе гребня и гроздей), ягодный показатель (число ягод в 100 граммах грозди).
Анатомическое строение ножки грозди и гребня сходно с анатомическим строением молодого побега. Хорошо развиты механические и проводящие ткани; из механических тканей — древесинные волокна (либриформ), клетки которой, являясь живыми, также выполняют запасающую функцию, и перицикловые тяжи. Из проводящих тканей сильно развит мягкий луб, снабжающий ягоды продуктами ассимиляции в период их роста и созревания. Плодоножка имеет подобное анатомическое строение.
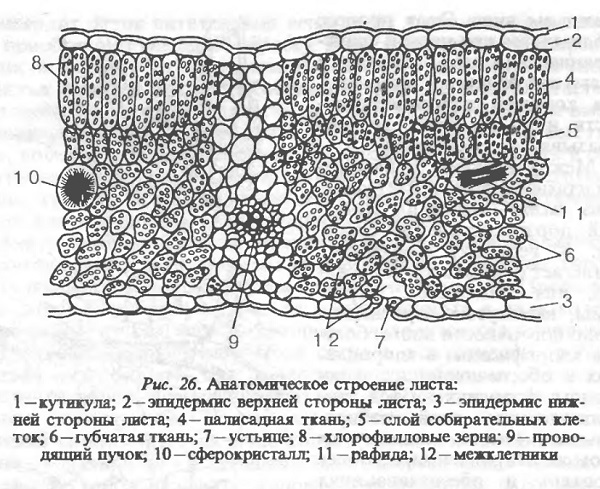
Ягода, расположенная на плодоножке (рис. 33), является сочным плодом винограда и самым важным органом, для получения которого возделывается виноградное растение, а также служит для защиты и распространения семян. Ягода формируется из завязи после цветения, опыления, оплодотворения и завязывания. В результате из яйцеклеток образуются зародыши, из семяпочек — семена, а из стенок и перегородки завязи — перикарпии (околоплодник), который является съедобной частью ягоды. Столбик и рыльце пестика отсыхают и остаются в виде тёмной точечки на вершине ягоды — пупочка. Мясистый перикарпий из стенок завязи формируется в основном за счёт роста, а не деления клеток. Рост клеток стенок завязи стимулируется эндогенными стимуляторами роста, образующимися в результате развития зародыша и семени, и обеспечивается притоком продуктов ассимиляции через проводящие пучки, проникающие в ягоду из плодоножки. Проводящие пучки, составляющую кисточку, разделяются в ягоде на три группы: периферические (питают периферическую часть ягоды), центральные (питают перегородку завязи) и питающие семена. В них хорошо развита флоэма и почки отсутствуют механические ткани. При полном созревании периферические проводящие пучки разрушаются и вместо них в мякоти образуются пустоты (аэрензима).
Ягода состоит из перикарпия и семян. Перикарпий ягоды состоит из трёх частей: экзокарпия (наружная эпидерма), мезокарпия (основная паренхима) и эндокарпия (внутренная эпидерма) (рис. 34). Экзокарпий состоит из одного наружного ряда клеток перикарпия, играющих защитную роль. Особое значение свойства этого слоя имеют для столовых сортов, определяя лёжкость и транспортабельность.
У молодых ягод некоторые клетки экзокарпия образуют устьица, которые впоследствии опробковевают и превращаются в чечевички. На наружной поверхности ягод образуется кутикула, состоящая из кутина и воска. Восковый налёт (пруин) защищает ягоду от потери воды, предохраняет от порчи, определяет транспортабельность лёжкость. Установлено также, что воск, входящий в состав налёта, дополнительно обогащает вина ароматическими веществами.
Эндокарпий представлен одним рядом мелких, толстостенных удлинённых клеток, составляющих внутренний эпидермис перикарпия и отделяющих его от сердечка — пустот, в которых располагаются семена. Если семена в гнёздах завязи не образуются, то перегородка её разрастается сильнее, заполняя пустоты.
Мезокарпий составляет среднюю и основную части перикарпия, он состоит из трёх зон. Наружная зона — гипокарпий — это 1…15 слоёв удлинённых толстостенных клеток, образующих вместе с экзокарпием и кутикулой кожицу ягоды. В клетках кожицы накапливаются химические вещества, обусловливающие окраску ягоды: антоксантины (хлорофилл, ксантофилл, каротин), флавоны (кветцетин, кверцитрин) и антоцианы, имеющие большое значени для качества винограда и особенно — вина. Доля кожицы от общей массы ягоды составляет 12…20% в зависимости от сорта и условий развития ягоды. Кроме красящих веществ, в клетках кожицы накапливаются дубильные и ароматические вещества, органические кислоты. Свойства кожицы определяют способность ягод к транспортировки, хранению, устойчивость к болезням и вредителям, растрескиванию. Различают следующие типы кожицы ягод винограда: тонкая, тонкая рвущаяся, тонкая, не отделяющаяся от мякоти, средней толщины, толстая и др. Следующие две зоны мезокарпия с разной толщины клеток образуют вместе с эндокарпием мякоть ягоды. Большая часть клеток мякоти имеет тонкие оболочки, крупные вакуали с соком. В вакуолярном соке содержатся сахара, органические кислоты, витамины и другие вещества; сусло в основном состоит из него. От толщины клеточных стенок мякоти, содержания протопектина и клетчатки, толщины проводящих пучков зависит консистенция мякоти. Это важнейший признак мякоти, определяющий направление использования продукции. В зависимости от сорта консистенция бывает сочной, слизистой, мясисто-сочной, мясистой, мясисто-хрустящей, хрустящей, хрящеватой. Для винных сортов желательна сочная мякоть, облегчающая извлечение сока. Столовые сорта винограда, особенно предназначенные для хранения и транспортировки, а также сорта для сушки должны иметь плотную мякоть. В период созревания мякоть составляет 75…85% массы ягод, а вакуолярный сок — свыше 99% мякоти. У большинства сортов мякоть не окрашенная, только у некоторых сортов клетки, а соответственно и сок имеют окраску — Саперави, Пти буше, Тентюрье.
Важнейшими признаками морфологической характеристики ягоды является окраска, величина и форма.
Окраске ягод винограда различных сортов присуще большое разнообразие от почти чёрной до молочно-белой. Обусловлено это различием в структуре пигментов, их количеством, глубиной залегания окрашенных слоёв клеток. Окраска ягод является важнейшим ампелографическим признаком, однако интенсивность её может изменяться в зависимости от условий произрастания. У представителей рода Витис различают следующиет типы окраски: тёмно-синяя, тёмно-красная, тёмно-коричнево-красная, розовая, тёмно-зелёная, жёлто-зелёная, зелёно-серая, коричневая, серая, зеленовато-коричневая и молочно-белая. Однако основными типами окраски являются жёлто-зелёная (белая), серая, розовая, красная и тёмно-синяя (чёрная).
Величина ягоды определяется сортом, условиями произрастания и выражается в единицах: линейных (мм), объёмных (см³) и массы (г). Масса ягоды колеблется от 1 до 10 г. Более крупные ягоды характерны для столовых сортов, самые мелкие — для коринок. Для описания величины ягод, имеющих неправильную форму, используют их объём, который определяется по объёму вытесненной ими воды — этот показатель изменяется от 0,5 до 11 см³. Чаще для характеристики величины ягоды используют величину среднего диаметра, который определяется как полусумма продольного и поперечного (в самой расширенной части) диаметров. По величине среднего диаметра различают следующие группы ягод: очень мелкие — меньше 8,0 мм, мелкие — 8,1…12,0; средние — 12,1…17,0; крупные — 17,1…25,0 и очень крупные — более 25,0 мм.
Для характеристики формы ягоды, наиболее стабильного признака сорта, используют индекс формы — отношение продольного диаметра к поперечному. По величине индекса формы различают следующие типы форм ягод: сплюснутая до 1,0; округлая — 1,0…1,1; овальная — 1,1…1,3; продолговатая — 1,3…1,6 и длинная более 1,6. Встречаются у отдельных сортов и другие формы ягод: яйцевидная, обратнояйцевидная, цилиндрическая, цилиндрическая с перехватом, серповидная и т. д. (рис. 35).
Химический состав ягод разнообразен. Наиболее важными компонентами являются сахара и органические кислоты. Количество сахаров и их состав колеблются в зависимости от года, условий вегетации, агротехники, сорта, степени зрелости от 12 до 30%. Из сахаров накапливаются, в основном, глюкоза и фруктоза. К периоду полного созревания в небольших количествах накапливаются и другие сахара — сахароза, галактоза, рамноза, рафиноза и некоторые другие. Содержание органических кислот в соке ягод в период созревания колеблется от 0,5 до 1,9%. Наиболее важные органические кислоты из группы нелетущих кислот (г/дм³) — яблочная (2…15), винная (2,0…7,0), лимонная (0,2…0,5), янтарная (0,1…0,3), щавелевая (до 0,15), слизевая (до 0,5), глюконовая (до 0,12), молочная (до 0,05) и др.; из группы летучих (мг/дм³) — муравьиная (20…50), уксусная (20…50), пропионовая, изомасляная, валериановая и др. Содержание сахаров и органических кислот и их соотношение определяют качество винограда и направление использования ягод. Для различных направлений использования и видов продуктов переработки установлены соответствующие кондиции по содержанию сахаров и кислот в соке ягод.
В соке ягод винограда присутствуют витамины Р, С, группы В (мезо-изозит — В₈, никотинамид — В5, рибофлавин — В2, фолиевая кислота — В9, тиамин — В1, пиридоксин — В6, пантотеновая кислота — В3) и др.
В ягодах накапливаются ароматические вещества, включающие группу химических соединений: спиртов, сложных эфиров, терпеноидов, карбонильных соедининий и др., входящих, в основном, в состав кожицы. Аромат обусловливается определённым сочетанием этих веществ. Установлено, что синтезируются ароматические вещества непосредственно в ягодах в конце фазы созревания.
Дубульные вещества накапливаются, в основном, в гребнях и семенах, небольшое количество — в кожице и мякоти ягод: в расчёте на сухую массу 4,7…11,3% и 0,6…2,5% соответственно. Дубильные вещества участвуют в формировании цвета, букета, типа вина, придают им вяжущий вкус.
Азотистые вещества, накапливающиеся в ягодах винограда, имеют различную природу. Общее их содержание в составе сусла — около 300…1930 мг/л в зависимости от места выращивания . В состав азотистых веществ винограда входят белковые вещества, пептоны, пептиды, аминокислоты, амиды, нитратные и аминные соединения. В процессе созревания ягод содержание азотистых веществ возрастает: аминного азота в 2 раза, белкового — в 10 раз; полипептидов — в 4 раза; доля аммонийных соединений снижается с 50% (от общего содержания азота в ягоде) до 15%, а при перезревании наблюдается дальнейшее резкое снижение. Следствием указанного является слабое брожение сусла вина, его цвета, букета, стабильности, имеют большое значение для качества коньяка.
Содержание пектиновых веществ в сусле колеблется от 0,5 до 2 г/дм³, причём их больше у красных сортов. Минеральные вещества в больших количествах накапливаются в гребнях, семенах. Содержание их в соке составляет 3…5 г/дм³, а в вине — 1,5…3 г/дм³, так как часть используется дрожжами, выпадает в осадок. Концентрация минеральных веществ в среднем составляет (мг/дм³): калия — 400…1800, кальция — 80…200, натрия — 20…200, магния — 50…150, железа — 1…20, меди — 0,1…5, алюминия — 1…2, цинка — 0,1…5, марганца 0,5…10 и др.
Совокупность веществ, входящих в состав ягод винограда, определяет вкус, аромат, привкусы, качество винограда и особенно вина. Вкус ягод определяется прежде всего содержанием сахаров и кислот и их соотношением. Ароматические вещества определяют специфические привкусы и ароматы: мускатный, паслёновый, земляничный и др.
Количество образовавшихся в ягоде семян зависит от числа оплодотворённых яйцеклеток и соответственно развившихся семяпочек. Если развиваются все семяпочки, в ягоде образуется 4 семени (при нарушении в строении завязи может и больше), оно обычно 2…3 семени. а в некоторых ягодах и у некоторых сортов семян и вовсе не образуется. У семенных сортов в пределах сорта, куста и грозди размер ягоды зависит от количества образовавшихся семян. Это связано с тем, что в процессе развития зародышей и семян синтезируются стимуляторы роста (гиббереллин), которые усиливают приток ассимилятов в растущую ягоду. С увеличением числа семян увеличиваются и размеры ягод. Семя стимулирует ту сторону ягоды, на которой оно располагается, а бессемянная сторона при этом отстаёт в развитии, что обусловливает асимметричность ягод (рис. 36). Стимулирующее влияние гиббереллинов на рост ягоды позволило разработать способы экзогенного их применения для увеличения размеров ягод бессемянных сортов.
У виноградного растения встречаются два типа бессемянности — партенокарпия и стеноспермокарпия. При партенокарпии ягоды развиваются из завязи без оплодотворения, они имеют мелкие размеры, округлую форму. Такой тип плодообразования называют горошением. Семена отсутствуют, внутри ягоды имеется только едва заметный след от деформировавшейся семяпочки. Партенокарпические ягоды характеризуются более ранним началом сахаранакопления и более высоким содержанием сахаров. Партенокарпия у винограда встречается в двух формах: облигатная — характерная для сортов группы коринок (Коринка чёрная, Коринка белая) и факультативная (функциональная) наблюдается у ряда семенных сортов. Факультативная или частичная, партенокарпия наблюдается у сортов, склонных к горошению, и у сортов с функционально-женским типом цветка при неблагоприятных для опыления и оплодотворения условиях. Вследствие этого снижается товарность гроздей и величина урожая. У ряда сортов отсутствует способность к образованию партенокарпических ягод: без оплодотворения цветки у них опадают.
Стеноспермокарпия (кишмишность) — вид плодообразования винограда при неполном оплодотворении. Существуют две точки зрения на механизм формирования бессемянных ягод кишмишного типа. По первому мнению, при образовании ягод такого типа происходит опыление и оплодотворение, а затем, по достижении развивающимися ягодами 2,5…3,0 мм в диаметре, зародыш погибает и прекращает рост семени, а вместо семян образуются рудименты. По второму мнению, оплодотворения не происходит, так как пыльцевые трубки не доходят до яйцеклетки. Для образования ягод этого типа обязательно опыление и наличие рудиментов семян. Ягоды имеют форму, свойственную сорту, а величина их зависит от степени развитости рудиментов. Облигатная форма такой бессемянности присуща кишмишным сортам (Кишмиш белый, Кишмиш чёрный и др.). Разработана классификация рудиментов семян (К. В. Смирнов). Встречается также факультативная форма стеноспермокарпии, что свойственно некоторым семенным сортам — эти сорта перспективны для использования в селекции на бессемянность в качестве материнских форм.
Семя развивается из семяпочки в результате двойного оплодотворения и формирования зародыша и является органом полового размножения винограда (рис. 37). Зрелое семя винограда имеет овальную форму, грушевидную форму, небольшие размеры (3…3,8×3…5 мм), состоит из тела семени и клювика. Для семени винограда характерна дорзивентральность. С брюшной стороны семени, обращённой внутрь ягоды, располагается шов, две впадины. Противоположная сторона называется спинной. На спинной стороне располагается халаза — кольцеобразная вдавленность, через которую в семя проникают сосудистые пучки, питающие его. Семена различных сортов отличаются по морфологическим признакам: величине, форме, длине клювика, по форме и расположению халазы.
Зрелое семя снаружи покрыто прочной кожурой, внутри находится эндосперм и зародыш, который располагается в клювике. Кожура семени состоит из внешнего и внутреннего покровов, каждый из которого состоит из трёх слоёв. Наружный слой внешнего покрова состоит из одного слоя вытянутых клеток. Под ними располагается промежуточный слой внешнего покрова, состоящий из нескольких слоёв мёртвых клеток, содержащий дубильные вещества и способный впитывать влагу и потому называемый губчатым. За промежуточным слоем располагается защитный слой внешнего покрова, состоящий из 2…6 слоёв каменистых клеток с толстыми одревесневевшими стенками и покрывается всё семя, за исключением халазы и клювика. Внутренний покров включает три слоя, каждый из которых состоит из одного слоя клеток. Эндосперм состоит из плотно прилегающих друг к другу клеток, богатых протоплазмой и запасными питательными веществами — белками, жирами, — которые используются при прорастании.
Соцветие винограда является частью зелёного побега, несущей на себе цветки, и представляет собой сложную кисть или метёлку. Состоит из центральной оси, осей следующих порядков ветвления (обычно до 3-го, реже до 4…5-го порядков), которые заканчиваются бутонами цветков. Нижняя часть центральной оси соцветия до первого ответвления составляет его ножку, посредством которой соцветие прикрепляется к узлу побега, его несущего. Бутоны цветков располагаются по три — тройчатками, из которых более развит центральный.
Соцветие имеет форму, в целом соответсвующую форме будущей грозди. Размеры соцветия, разветвлённость осей, количество цветков зависят от сорта и условий формирования. У сортов, имеющих соцветия, образуется 50…100 цветков в одном соцветии, у сортов с крупными соцветиями — 700…1200 и более.
На одном плодоносном побеге образуется 1…3 соцветия (очень редко до 7…13). Соцветия развиваются на самых нижних симподиальных узлах, напротив места прикрепления листа. Выше последнего соцветия развиваются усики. Выше усика соцветия не образуются. Соцветия могут образовываться также на пасынках, очень редко на волчковых побегах. Все элементы соцветия — главная ось, боковые ответвления, бутоны — имеют зелёную окраску, иногда с красноватым оттенком (характерен для некоторых сортов). Если на плодоносном побеге образуется несколько соцветий, то самые крупные из них — нижние, затем последовательно размеры соцветий уменьшаются.
Закладываются соцветия в предшествующий своему появлению год в центральных почках зимующих глазков. Размеры зачаточного соцветия ко времени вступлния в покой почек и степень его дифференциации зависят от сорта и условий морфогенеза глазков и определяют во многом величину соцветий, образующихся из них в последующую вегетацию. Чем интенсивнее развивается центральная почка глазка, тем крупнее и дифференцированнее зачаточные соцветия. Самые крупные и дифференцированные зачаточные соцветия находятся в глазках, расположенных в средней части однолетнего побега. Если на зачаточном побеге центральной почки зимующего глазка, располагаются несколько соцветий, то их размеры и степень дифференциации уменьшаются а акропетальном порядке. Более развитые по размеру и дифференциации соцветия формируются на полноценных побегах. Количество и развитость эмбриональных соцветий зависят от силы куста, погодно-климатических, почвенных условий, агротехники. Использование приёмов, обеспечивающих оптимальное соотношение между ростом и плодоношением (прищипывание и чеканка побегов, обработка регуляторами роста, подкормка, ориентация растущих побегов в пространстве и др.), может стимулировать закладку и развитие зачаточных соцветий.
Весной с началом развития почек возобновляется рост и дифференциация зачаточных соцветий: формирование примордиев в цветки и образование элементов цветка. Условия этого периода играют большую роль в реализации эмбриональной плодоносности почек винограда. При благоприятных условиях формируются соцветия полного развития. Если условия неблагоприятные, то происходит частичная или полная редукция зачаточных соцветий в усики.
При частичной редукции образуются переходные от соцветия к усику формы с разной степенью развитости генеративной и вегетативной частей и происходит также определённая саморегуляция растением уровня нагрузки элементами плодоношения. Этот период профессор А. М. Негруль назвал «вторым критическим периодом в формировании генеративных органов». На процесс реализации эмбриональной плодоносности почек влияют такие факторы, как погодные условия, удобрение, уровень влагообеспеченности, уход. Чрезмерно быстрое нарастание температуры весной и быстрое распускание почек приводит к редукции элементов плодоношения и наоборот — постепенность этого процесса способствует более полной реализации эмбриональной плодоносности. При быстром нарастании температуры воздуха почва не успевает прогреться и интенсивность роста корней не соответствует интенсивности роста надземных органов, что приводит к дефициту питания распускающихся почек. В этой связи при выращивании винограда в защищённом грунте необходимо пред повышением температуры полить грунт в теплице тёплой водой. Это обеспечивает оптимальную последовательность в развитии корней и почек. Приёмы, задерживающие быстрое распускание почек винограда весной в открытом грунте, способствуют полной реализации эмбриональной плодоносности, формированию большего количества соцветий полного развития.
С началом распускания почек и роста побегов происходит и интенсивный рост соцветий (рис. 27). Сперва соцветия прикрыты на растущей верхушке нижерасположенными листьями. По мере роста побега и осей соцветия они высвобождаются из листьев и становятся видными при длине побега 15…20 см. Рост соцветий постепенно достигает максимума, затем снижается и почти прекращается к началу цветения. В период массового цветения рост осей соцветия возобновляется.
Зона максимальной интенсивности роста главной и боковых осей соцветия перемещается в акропетальном порядке, постепенно ослабляясь. Вследствие этого соцветие приобретает в целом конусовидную форму. Развитость элементов соцветия определяет размер будущей грозди.
Иногда в развитии соцветий наблюдаются уродства (фасциации), которые выражаются в образовании уродливых, широких и плоских осей соцветия, групп бутонов с укороченными цветоножками.
Анатомическое строение осей соцветия подобно таковому молодого побега и соответствует выполняемым фунциям: хорошо развита проводящая система (особенно флоэма) и механические ткани (колленхима). Так же как и для побега, для них характерна дорзевентральность в развитии тканей. Хорошо развита сердцевина.
Цветки винограда мелкие, невзрачные, желтовато-зелёной окраски пятерного типа и имеют формулу Ca(5)Co(5)A(5)C(2) (рис. 28). Цветок винограда прикрепляется к оси соцветия при помощи тонких цветоножек. Цветоножка после завязывания ягоды превращается в плодоножку. Через неё проходят сосудисто-проводящие пучки. В верхней части цветоножка расширяется и образует цветоложе, которое затем развивается в подушечку ягоды.
Органы цветка располагаются симметрично, кругами (рис. 29). Наружный круг состоит из пяти недоразвитых сросшихся чашелистиков, имеющих вид слабовыступающих тупых зубчиков, и формирует чашечку цветка. Второй круг состоит из пяти сросшихся краями лепестков зелёной окраски. Они образуют венчик в виде колпачка, покрывающего цветок сверху и защищающего его внутренние части. Тритий круг образует андроцей, который состоит из пяти тычинок, а тычинка — из пыльника, связника и тычиночной нити. Длина тычиночной нити и её форма зависят от типа цветка, сорта. У обоеполых цветков, а также у сортов с функционально-мужским типом цветка тычиночные нити длинные, прямостоячие, а у сортов с функционально-женским типом цветка — короткие, загнутые вниз. Пыльник состоит из пыльцевых мешков, попарно соединённых широким связником, который является продолжением тычиночной нити. В пыльниках образуется пыльца, состоящая из пыльцевых зёрен. Пыльцевые зёрна мелкие, овальной формы, бочкообразные, длиной 18…50 мкм. Сформированное фертильное пыльцевое зерно винограда включает генеративную клетку, вегетативное ядро и имеет поры, покрытые двойной оболочкой — интиной и экзиной. Пыльца, попавшая на рыльце пестика или помещённая в искусственную питательную среду, набухает, приобретает сферическую форму, очень быстро образует в покровах выпячивание, из которого образуется пыльцевая трубка. Оптимальная температура для прорастания пыльцы 20…30º С, при 13…14º С пыльца не прорастает, а выше 35º С прорастание в значительной степени тормозится.
Четвёртый круг цветка представлен гинецеем синкарпного (сростнолистного) типа, состоящим из двух сросшихся плодолистиков, образующих пестик. Пестик, расположенный в центре цветка, имеет грушевидную форму и включает завязь, столбик и рыльце. Форма пестика, его размеры являются характерными признаками сортов винограда.
Завязь виноградного цветка нижняя, двухгнёздная, состоит из двух сросшихся плодолистиков. Гнёзда разделены перегородкой, и в каждом из них закладывается по две семяпочки. Из завязи формируется плод винограда. Семяпочка винограда базальная, формируется у основания завязи (рис. 30). Снаружи она покрыта наружным и внутренним интегументом, в которм имеется микропиле — канал, через который пыльцевая трубка проникает в семяпочку. Внутри семяпочки — многоклеточное ядро нуцеллус, который включает в себя зародышевый мешок. В зародышевом мешке оплодотворяется яйцеклетка, формируется зародыш и эндосперм. Семяпочка соединяется с основанием завязи посредством семяпочки, через которую проникает в семяпочку проводящий пучок, доходящий до халазы, расположенной вверху семяпочки. У некоторых цветков встречаются отклонения: количество гнёзд и семяпочек в них увеличивается, тогда в ягодах появляется более четырёх семян.
Столбик расположен в верхней части пестика, имеет разную длину и форму в зависимости от сорта. Через столбик проходят проводящие пучки, по которым питательные вещества поступают к рыльцу пестика. Внутренняя часть столбика состоит из рыхлой ткани, облегчающей проникновение пыльцевой трубки в завязь.
Рыльце пестика воспринимает пыльцу и обеспечивает условия для её прорастания. Оно имеет блюдцеобразную форму с приподнятыми краями. Во время цветения на рыльце пестика выделяется липкая жидкость для прилипания пыльцы к рыльцу. У основания завязи на подпестичном диске, по кругу чередуясь с тычинками, образуются 5 нектарников, выделяющих эфирные масла с приятным ароматом. Нектарники имеют различную форму, размеры и их морфологические признаки используются при описании сортов.
В зависимости от наличия в цветке мужской и женской части и степени их развития различают следующие типы цветков: истинно-женский, мужской, обоеполый, функционально-мужской, функционально-женский (рис. 31). Три последних типа являются для винограда основными. Истинно-мужской цветок у винограда не обнаружен, но отмечено образование истинно-женских цветков у растений сорта Мурведр.
Цветок обоеполого типа, описанный выше, характерен для подавляющего большинства культурных сортов. У цветков этого типа — прямостоячие длинные тычинки с фертильной пылцой.
Функционально-мужской тип отличается тем, что цветок имеет недоразвитый пестик, а мужская часть развита нормально и формирует фертильную пыльцу. Редукция пестика у цветков может быть выражена по-разному: от маленького бугорка в центре цветоложа до довольно крупной завязи. Общим для них является недоразвитость семяпочке, развитие которых прерывается на различных этапах. Этот тип цветка не способен формировать плод и характерен для сортов-подвоев и встречается у особей дикого винограда.
Цветок функционально-женского типа имеет нормально развитый пестик, способный воспринимать пыльцу и формировать ягоду, и недоразвитую мужскую часть. Тычинки недоразвитые, короче пестика и загнуты вниз или закручены. Пыльца, образующаяся в пыльниках, недоразвитая, нежизнеспособная — организационно-стерильная. Пыльцевые зёрна имеют по сравнению с фертильными более вытянутую форму с заострёнными концами, поры почти отсутствуют. Такой тип стерильности обусловлен характерными для этих цветков дегенерационными процессами в период формирования тычинок. Эта пыльца не способна к прорастанию и оплодотворению, однако стимулирует образование бессемянных мелких ягод при попадании на рыльце. Для формирования ягод из таких цветков требуется жизнеспособная пыльца с функционально-мужских или обоеполых цветков. Функционально-женский тип цветка имеют некоторые культурные сорта (Нимранг, Чауш белый, Пухляковский, Мадлен Анжевин и др.) и особи диких форм. Дикий виноград V. vinifera ssp. silvestris — двудомное растение. У некоторых представителей семейства Vitaceae выражена полигамность: наличие на одном и том же растении обоеполых и однополых цветков.
В строении и развитии цветков винограда наблюдаются аномалии, т. е. отклонения от нормального роста и развития. У сортов с обоеполым или мужским типом цветка под влиянием неблагоприятных условий образуется случайно-стерильная пыльца, недоразвитая, деформированная и нежизнеспособная. Образование большого количества такой пыльцы ухудшает опыление и приводит к снижению урожая. Отмечаются также аномалии в виде махровости, раскрытия цветка розеткой. Махровость проявляется в превращении тычинок, пестика, нектаринов в лепестки и т. д. Отмечены случаи слияния двух цветков в один сложный, двух-трёх тычинок в одну, образование многогнёздных завязей, большего числа семяпочек и др. Причины этих явлений — заражение вирусами, бактериями, грибами, воздействие ионизирующих изделий, химических мутагенов, мутации.
Перед началом цветения в месте прикрепления лепестков венчика к цветоложу образуется рыхлая отделяющая ткань. Затем лепестки отделяются от цветоложа, разъединяются с нижней стороны по швам и под давлением выпрямляющихся тычинок сбрасываются в виде колпачка (рис. 32). У обоеполых цветков одновременно растрескиваются пыльники и пыльца попадает на рыльце пестика, на котором к этому времени выделяется секреторная жидкость.
Обоеполые цветки опыляются или собственной пыльцой (в большинстве случаев), или пыльцой с цветков этого же соцветия, или других соцветий этого же куста. Сорта, имеющие функционально-женский тип цветка, опыляются пыльцой других растений, имеющих обоеполый тип цветка. Для этих сортов осуществляют смешанные посадки: на каждые 1…2 ряда сорта с функционально-женским типом цветка высаживают 1…2 ряда сорта-опылителя, имеющего обоеполый тип цветка и такие же сроки цветения.
Виноград — растение анемофилное, т. е. пыльца переносится ветром. В тех случаях, когда пыльники лопаются до опадения колпачка, происходит самоопыление обоеполых цветков при нераскрывшемся цветке — клейстогамия. Причиной являются неблагоприятные погодные условия (засуха, низкая температура) в период, предшествующий цветению. Существуют сорта, склонные к клейстогамии. Клейстогамия отмечена также при обработке некоторыми регуляторами роста. В результате такого опыления завязывается ягода, в процессе её роста лепестки открываются друг от друга и раскрываются в виде звёздочки.
Рыльце пестика восприимчиво к приёму пыльцы 4…6 дней с момента опадения венчика. Если нет опыления, цветки отсыхают и опадают.
Пыльцевые зёрна, попавшие на рыльце пестика, набухают, прорастают, образуюя пыльцевую трубку. Пыльцевые трубки проникают через столбики к завязям и достигают семяпочки через 15…24 ч после начала прорастания. Через пыльцевую трубку в семяпочку проникают спермии, и происходит процесс двойного оплодотворения. Процесс оплодотворения осуществляется при температуре 14…15º С. При наличии физиологических барьеров для оплодотворения (несовместимость, самостерильность, невосприимчивость) наблюдается образование укороченных или удлинённых пыльцевых трубок с утолщёнными концами, их рост в сторону от завязи.
После оплодотворения яйцеклетка остаётся на 10…20 дней в состоянии покоя, затем начинается формирование зародыша. Оплодотворённое вторичное ядро сразу начинает делиться, формируя ядра эндосперма.
Усик выполняет механическую функцию прикрепления растения к опоре. Так же, как и соцветие, возникает из отклонившейся точки роста в симподиальных узлах, т. е. является видоизменённым побегом. Усики, образующиеся на нижних (3…14-м) узлах побегов, закладываются в процессе морфогенеза почек в год, предшествующий их развитию, остальные закладываются и развиваются в текущем году. На бесплодных побегах усики образуются, начиная с 3…4-го узла, напротив места прикрепления листа, а на плодоносных — выше узлов, несущих соцветия.
Усик растёт по симподиальному типу. Усики бывают простые (неразветвлённые) и разветвлённые. Простые усики характерны для представителей подрода Muscadinia, разветвлённые — для представителей подрода Euvitis. На каждом узле усика, где происходит ветвление, образуется недоразвитый лист в виде чешуйки.
В редких случаях вместо усика образуется внепазушний побег, это часто наблюдается у сорта Альбурла. Существуют также переходные формы от усика к соцветию.
Во время роста побега усики появляются на верхушке побега. Кончик усика растёт и совершает нутационные круговые движения из-за неравномерного роста клеток на разных сторонах. Если при этом кончик усика встречает опору, он испытывает односторонне раздражение, воспринимаемое особыми клетками эпидермиса (гаптотропизм), вследствие чего усик усиливает рост и кончик его обвивает вокруг опоры, а нижняя часть изгибается в виде спирали, подтягивая побег к опоре. Затем ткани усика быстро одревесневают, прочно закрепляя побег на опоре. Если усик не встречается с опорок, он остаётся травянистым, отсыхает и опадает.
У некоторых представителей семейства Vitaceae (род Partenocissus) усики заканчиваются присосками, которые позволяют закрепляться им на гладкой поверхности.
При выращивании винограда в культуре на опорах усик не имеет значения как орган, выполняющий механическую функцию, более того, он затрудняет выполнение обрезки, съём побегов со шпалеры. Поэтому в некоторых частных случаях практикуют удаление усиков: в защищённом грунте, на маточниках.
Лист — это боковой вегетативный орган виноградного растения, выполняющий ряд важнейших функций. В листе происходит процесс фотосинтеза, т. е. преобразование неоганических веществ в органические. В результате транспирации, осуществляемой листьями, исключается перегрев органов, развивается сосущая сила, обеспечивающая непрерывное поглощение и передвижение воды и питательных веществ в растении. Благодаря дыханию в листьях высвобождается энергия, необходимая для ряда других процессов метаболизма. Через листья осуществляется интенсивный газообмен между растением и окружающей средой.
По происхождению различают листья двух типов: семядольные листья и листья, образующиеся на конусе нарастания побега (настоящие). Семядольные листья образуются только у сеянца винограда. Они закладываются на материнском растении в процессе развития зародыша семени. Семядоли содержат запасы питательных веществ, используемых зародышем при прорастании. При прорастании семядоли выносятся на поверхность субстрата, растут, зеленеют и выполняют все основные функции листа. Семядольные листья сильно различаются по форме, размерам и внутреннему строению от обычных листьев. Все последующие листья сеянца, как и все другие листья винограда, закладываются в результате деятельности конуса нарастания.
Первые 3…14 листьев саженца также закладываются в процессе органогенеза почки на материнском растении, из которой развивается надземная часть. Специфика в развитии этих, так же как и нижних 9…14 листьев, плодоносных и бесплодных побегов заключается в том, что они перезимовывают в виде зачатка в зимующем глазке. Остальные листья основных побегов и пасынков закладываются и развиваются в текущем году.
Листовой бугорок, отчленившийся на конусе нарастания побега, вытягивается, дифференцируется и формирует листовую пластинку, черешок и два пленчатых сидячих бледно-зелёной окраски прилистника. Прилистники появляются на верхушке побега, располагаются с двух сторон в месте прикрепления черешка к побегу и прикрывают верхушку, выполняя защитную функцию. По мере удаления узла, их несущего, от верхушки прилистники постепенно отсыхают и отпадают.
Лист винограда простой, состоит из черешка и листовой пластинки (рис. 20). Черешком называют суженная стеблевидная часть листа, он выполняет механическую функцию поддержания листовой пластинки оптимальной ориентации её по отношению к свету. По проводящей системе черешка осуществляется отток ассимилятов из листовой пластинки и снабжение её водой и питательными веществами. В тканях черешка кратковременно сохраняются питательные вещества. Черешок у листа винограда, имеет зелёную или, осенью, различной интенсивности красноватую окраску, опушение. Нижняя часть черешка расширена и в месте прикрепления обхватывает побег с трёх сторон. По мере удаления от побега черешок приобретает цилиндрическую форму. В месте прикрепления к листу черешок расширяется и образует со своей верхней стороны углубление в виде желобка. Входя в листовую пластинку, черешок разветвляется и формирует пять основных жилок листа, которые, в свою очередь разветвляясь, образуют густую сеть жилкования. Длина черешка зависит от сорта и условий выращивания. У затенённых листьев более длинные черешки.
Листовая пластинка у винограда крупная, расширенная, плоская, по краям зубчатая (рис. 21), разнообразной формы, редко цельная, чаще лопастная.
Форма листовой пластинки в зависимости от сорта бывает округлой, яйцевидной, сердцевидной, клиновидной, почковидной. По рассечённости: цельные, слабо-, средне-, глубокорассечёнными (рис. 22), трёх- или пятилопастные (очень редко семилопастные).
Лопасти листа отделяются друг от друга боковыми вырезками и черешковой выемкой. Боковые вырезки (рис. 23) и черешковая выемка (рис. 24) у различных сортов развиваются по глубине, открытости, виду просвета, характеру дна.
Листовая пластинка состоит из мякоти и жилок. Жилки выступают в виде рёбер с нижней, а иногда и с верхней стороны листа. Они выполняют механическую и транспортную функции. В центре располагается главная жилка, которая делит листовую пластинку на две половины. Боковые жилки отходят от места прикрепления черешка к листовой пластинке под определённым углом по отношению друг к другу, называемым углом жилкования. От основных жилок отходят жилки следующего порядка и т. д. Жилки последующего порядка заканчиваются в мякоти листа или входят в зубчики по краям листа. Тип зубчиков по краям листа и на вершинах лопастей используется для ампелографического описания сортов.
Поверхность листовой пластинки винограда бывает гладкой, сетчато-морщинистой, пузырчатой. Она бывает ровной или с изгибающимися вверх или вниз краями. Молодые листья, расположенные на верхушке побега, опушенные сильнее, чем взрослые листья, и имеют окраску различный оттенков. Пасынки взрослых листьев винограда могут быть голыми или опушенными, как правило, с нижней стороны. Опушение листьев бывает щетинистым, паутинистым или смешанным (рис. 25).
Окраска листьев изменяется от светло-зелёной до тёмно-зелёной в зависимости от сорта и условий произрастания. Осенью перед листопадом листья приобретают характерную осеннюю окраску: листья сортов с белыми и розовыми ягодами становятся жёлтыми, а с чёрными — красными.
Морфологические признаки листьев изменяются в зависимости от сорта, возраста листа, узла, на котором они располагаются. Наиболее типичные для сорта листья располагаются на 8…12-м узле.
Листья винограда относятся к органам с ограниченным ростом, т. е. растёт в течение 5…6 недель, достигая определённого размера, после чего рост его прекращается. По мере роста усиливается процесс ассимиляции, изменяется химический состав листа. Осенью активность ассимиляци падает, хлорофилл разрушается, происходит отток питательных веществ из листа в стебель, листья приобретают осеннюю окраску. В месте прикрепления черешка побега остаётся листовой след. Если поверхность листового следа гладкая и имеет коричневую окраску, это значит, что листья кончили вегетацию, побеги вызрели нормально. Шероховатая поверхность и серо-грязная или чёрная окраска листового следа свидетельствуют о том, что листьч не завершили вегетацию вследствие затягивания её или заморозков и побеги плохо вызрели.
Анатомическое строение черешка, жилок и листовой пластинки соответствует выполняемым ими функциям. В черешке хорошо развиты проводящая система и механическая ткань. Проводящая система черешка состоит из расположенных в виде колец проводящих пучков, между которых находятся сердцевинные лучи. Большое количество пучков, сосудов и ситовидных трубок создаёт возможности для интенсивного снабжения листовой пластинки водой, элементами питания и быстрого оттока продуктов ассимиляции. Успешному выполнению механических функций способствует наличие кольца первичной механической ткани — колленхимы — и перецикловых склеринхимных волокон. В черешке хорошо развиты коровая и древесинная парехимы, создающие возможность для временного хранения запасных питательных веществ при их передаче в лист или в стебель. Снаружи черешок, так же как и жилки, покрыт эпидермисом и кутикулой. Главные жилки, которые образуются черешком при входе в листовую пластинку, имеют подобное же строение, только меньшее количество проводящих пучков. В жилках последующих порядков, при сохранении общей анатомической структуры, последовательно уменьшается размер и количество проводящих пучков.
В межжилковом пространстве анатомическое строение листа имеет обычную структуру (рис. 26). С верхней и нижней стороны лист покрыт эпидермисом, состоящим из одного слоя плотно прилегающих друг к другу клеток, снаружи которых образуется кутикула. Эпидермальные клетки черешка и листовой пластинки, особенно с нижней её стороны, образуют выросты (волоски), составляющие опушение различных типов. На нижней стороне листовой пластинки (на верхней очень редко) клетки эпидермиса образуют устьица, обеспечивающие регулируемый обмен с окружающей средой. Их количество на единицу площади листа в зависимости от сорта, возраста листа, места на побеге, условий формирования (влажность, освещённость и др.) колеблется от 120 до 200 шт. на 1 мм². На концах зубчиков листьев формируются водные устьица (гидатоды), которые способны выделять воду в жидком виде. Этот процесс наблюдается при низкой транспирации, высокой влажности (часто встречается в теплицах), при хорошей влагообеспеченности и на молодых листьях и называется гутацией.
Между верхним и нижним эпидермисом располагается мякоть листа — мезофилл, имеющий дорзивентральное строение. К верхнему эпидермису прилегает слой клеток палисадной, или столбчатой, паренхимы, вытянутых перпендикулярно поверхности листа, богатых хлорофиллом в хлоропластах и обеспечивающих, в основном, фотосинтез. Под ним располагается слой собирательных клеток, вытянутых в том же направлении, но более коротких и обеспечивающих передачу продуктов ассимиляции проводящей системе жилок, которые своими окончаниями прилегают к ним.
Под этим слоем, примыкая к нижнему эпидермису, располагается губчатая ткань, состоящая из 4…7 слоёв неправильной округлой формы клеток с крупными межклетниками. Клетки губчатой ткани содержат небольшое количество хлоропластов и выполняют функцию газообмена и также передают ассимиляты в проводящую систему жилок.
Листья винограда характеризуются очень высоким уровнем важнейших процессов жинеобеспечения растительного организма — фотосинтеза, транспирации, дыхания. В результате деятельности отдельных листьев формируется совокупная биологическая и хозяйственная продуктивность побега, виноградного куста и насаждения. Отдельные листья при этом функционируют в условиях не характерного для естественно произрастающего растения, а создаваемого искусственного ценоза.
Листовой поверхностью называется суммарная площадь листьев побега, куста, ряда и виноградника. Величина листовой поверхности, её структура, условия её функционирования определяют величину биологического и хозяйственного урожая, качество продукции. Существуют различные методы определения величины листовой поверхности: весовой, планиметрический, рассчётный (ампелометрический). Сущность наиболее распространённого в виноградарстве ампелометрического метода заключается в том, что площадь листа рассматривается как площадь круга с диаметром, равным условному диаметру листа (от верхнего зубца на центральной лопасти до наиболее удалённого зубца нижней боковой лопасти). Затем вносится поправочный коэффициент на рассечённость листовой пластинки, который составляет для целых листьев 1,25; трёхлопастных — 1,27; для пятилопастных слаборассечённых — 1,3 и сильнорассечённых — 1,35.
Листовая поверхность побега достигает 16…18 дм² и больше, куста — 11…14 м², гектара виноградника — 35…40 тыс. м². Наиболее существенно на этот показатель влияет нагрузка побегами, форма, уход, влагообеспеченность и др.
Однако интенсивность фотосинтетической деятельности, интенсивность и направленность других важных процессов метаболизма определяются архитектурой куста — расположением листовой поверхности в пространстве и развитием её во времени (фотосинтетический потенциал). При оценке кроны куста с точки зрения поглощения ФАР её моделируют как призму, цилиндр и другие геометрические фигуры. Естественно, что листья, расположенные на разных ярусах, различных слоях, получают разную величину ФАР и характеризуются разной продуктивностью. Индекс листовой поверхности рассчитывается как отношение односторонней площади листьев к площади поверхности земли, которую эта листовая масса занимает. Показатель является весьма важным для характеристики величины листовой поверхности, условий функционирования листьев. Для этих же целей используют показатель плотности площади листьев — отношение площади листьев к объёму кроны, в которой они располагаются (м²/м³). Необходимо иметь в виду, что оптимальные величины этих показателей для крон различной архитектуры (штамбовые, бесштамбовые, способ ведения прироста, плотность посадки) неодинаковы.
Важнейшим показателем, характеризующем листовую поверхность винограда, является фотосинтетический потенциал — сумма ежедневных площадей листьев куста, ряда или насаждения за период формирования урожая или часть этого периода. Различным величинам фотосинтетического потенциала каждого сорта соответствует разная величина биологического и хозяйственного урожая. Так, для получения 100 ц/га урожая гроздей сорта Ркацители необходима расчётная величина фотосинтетического потенциала составляет 1,26…1,76 ∙ 10⁶ м² ∙ день, а для получения 150 ц/га — 1,83…2,64 ∙ 10⁶ м² ∙ день.
Листовая поверхность куста и насаждения нарастает постепенно: максимальная величина её достигается к началу созревания, к началу цветения — 15…30% от неё, а через 20…25 дней после цветения — 60…65% от максимальной.
Интенсивность фотосинтеза оценивается через количество углекислоты, ассимилированной листьями. Этот показатель фотосинтетической деятельности виноградного растения зависит от ряда внешних и внутренних факторов: освещённости, температуры, влагообеспеченности, поступления питательных веществ и углекислоты, сорта, вида, возраста листьев, возраста кустов, агротехнических приёмов и др.
Фотосинтез возможен только на свету.Компенсационная точка светового фактора, т. е. величина освещённости, когда количество углекислоты, поглощаемой при фотосинтезе и выделяемой при дыхании, одинаково, в зависимости от сорта и вида колеблется в пределах 300…1500 лк. Наиболее низких уровень компенсационной точки характерен для вида V. riparia. По мере увеличения освещённости увеличивается и интенсивность фотосинтеза — эта зависимость имеет вид гиперболической кривой. Световая насыщенность фотосинтеза у винограда разных видов наступает при разных уровнях освещённости, начиная с 30…40 тыс. лк. Несмотря на высокие показатели светового насыщения, необходимо иметь в виду, что по мере удаления листьев от поверхности вглубь кроны значительно снижается освещённость. У затенённых листьев внутри кроны при загущении не только слабая интенсивность фотосинтеза, но они тратят большее количество пластических веществ на дыхание, особенно при повышенных температурах. Поэтому важны приёмы, регулирующие световой режим виноградника.
Оптимальной температурой для фотосинтеза является 28…32º С. При температуре выше 35º С и ниже 6º С фотосинтез прекращается.
Важнейшим фактором фотосинтеза является и водный режим. При недостатке влаги интенсивность ассимиляции резко падает. В засушливые годы в растении плохо накапливаются запасные и защитные вещества, медленно созревают ягоды, плохо вызревают побеги, в результате снижается морозо- и зимостойкость растений. При относительной влажности воздуха ниже 50% и влажности почвы ниже 45% ППВ существенно снижается интенсивность фотосинтеза.
На интенсивность фотосинтеза влияет концентрация углекислоты в воздухе. С повышением концентрации СО2 до 0,1% происходит крутой подъём интенсивности фотосинтеза, дальнейшее её повышение, однако, менее эффективно.Отмечено также, что повышение концентрации СО2 резко усиливает эффективность использования влаги. Внесение в почву виноградника органических удобрений способствует увеличению концентрации СО2 в приземном слое воздуха и стимулирует фотосинтез.
На интенсивность фотосинтеза влияют и элементы минерального питания. Недостаток или избыток некоторых из них может тормозить фотосинтез.
Для сохранения интенсивности фотосинтеза на высоком уровне необходим постоянный отток продуктов ассимиляции из листьев. Поэтому на кустах с увеличением нагрузки урожаем интенсивность фотосинтеза повышается. И, наоборот, с увеличением листовой поверхности интенсивность фотосинтеза снижается. Поэтому листья плодоносных побегов отличаются более высоким уровнем фотосинтеза.
В фотосинтезе листьев винограда существует суточная и сезонная динамика. В более тёплых районах виноградарства в течение дня наблюдается два пика интенсивности фотосинтеза: первый пик — в полуденные часы, затем — спад в середине дня и второй пик — после полудня. Установлено, что для листьев разных ярусов пик интенсивности наблюдается в разное время дня.
Фотосинтетическая активность листьев разных ярусов на побеге меняется в течение вегетации, зона максимальной интенсивности фотосинтеза смещается в акропетальном порядке. Меняется и направление перемещения ассимилятов от листьев разных ярусов. Количество листьев, работающих для нисходящего оттока ассимилятов, увеличивается: перед цветением один лист, расположенный выше соцветия, направляет ассимиляты вниз, после цветения — четыре, в начале созревания ягод — десять и т. д. К периоду созревания нижние 3…4 листа практически прекращают снабжать грозди углеводами, вместе с тем их наличие ухудшает микроклимат в зоне гроздей. Поэтому рекомендуется в этот период удалять.
Благодаря фотосинтезу нижних 8…10 листьев формируется масса гроздей, а на вышерасположенных лежит накопление сахаров и формирование качества. Из этого можно сделать вывод: для накопления сахаров имеют значения листья пасынков. Всё изложенное необходимо учитывать при проведении операций с зелёными частями куста.
Дыхание — важнейший процесс метаболизма, в ходе которого сложные органические соединения окисляются, в результате чего выделяется свободная энергия, используемая для роста, развития и т. д. В результате дыхания образуется углекислый газ и вода. Интенсивность дыхания зависит от ряда внешних и внутренних факторов. С повышением температуры интенсивность дыхания листьев возрастает, максимальная её величина наблюдается при 40º С. При температуре, повреждающей протоплазму, дыхание снижается. Интенсивность дыхания молодых листьев в несколько раз выше, чем взрослых. Установлены также изменения в интенсивности дыхания листьев в течение вегетации: в начале вегетации она нарастает, достигает максимума в период массового цветения, затем снижается к листопаду. Средняя величина интенсивности дыхания для листьев винограда составляет 1,5…2,5 мг/дм²∙ч.
Одной из важнейших функций листа является транспирация — процесс испарения воды. Вода испаряется в основном через устьица (устичная транспирация), а в небольшом объёме — через кутикулу (кутикулярная транспирация). Потеря воды клетками листа приводит к возникновению сосущей силы, которая, передаваясь от клетки к клетке, затем — к сосудам древесины и далее к корням, обеспечивает непрерывное поглощение и подачу воды и питательных веществ в листья. Другая важная роль транспирации — предотвращение перегрева листьев, а также других органов. Поступающая в листья вода также участвует в процессе фотосинтеза. Вместе с тем чрезмерно интенсивная транспирация при недостатке воды в почве приводит к завяданию листьев.
Виноградному растению присуща высокая интенсивность транспирации. Так, в опытах с сортом Рислинг установлено, что листья площадью 1 м² транспирируют 1,9…2,0 г воды в минуту. Один куст, имеющий 150…200 листьев, при температуре 24º С испаряет 1,0…1,5 л, а виноградник площадью 1 га с общей листовой поверхностью 30 000 м² — 30 т воды в сутки. Транспирационный коэффициент винограда составляет 120…400 л на 1 кг сухого вещества. Из общего количества поглощаемой растением винограда воды на формирование биомассы уходит 0,2%, остальное расходуется на транспирацию.
Транспирация имеет определённую суточную динамику: в утренние часы интенсивность её повышается и достигает максимума в околополуденные часы. Одним из важнейших факторов, определяющих величину транспирации, являются температура и влажность воздуха, а также влажность воздуха. Резко повышает транспирацию сухой воздух, ветер. На 2…7-ой день после полива у орошаемых кустов транспирация в 1,5…2,5 раза больше, чем у неорошаемых кустов. Установлены видовые и сортовые различия в интенсивности фотосинтеза: V. vinifera обладает самой большой транспирационной активностью, а V. riparia — самой низкой.
Виноградное растение способно регулировать интенсивность транспирации благодаря способности устьиц активно закрываться. При недостатке влаги устьица закрываются, транспирация ограничивается. Одновременно снижается интенсивность ассимиляции, так как уменьшается поступление углекислоты из атмосферы.
Почка представляет собой зачаточное состояние одного побега. У винограда почки выполняют ряд функций: роста, возобновления роста, вегетативного размножения. В почках закладываются эмбриональные генеративные органы и происходит их зачаточная дифференциация.
Почка состоит из конуса нарастания, обладающего меристематической активностью; зачаточного стебля, состоящего из зачаточных узлов и междоузлий. На зачаточных узлах располагаются зачатки листьев, усиков, соцветий, почек следующего порядка. Главной, неотъемлемой составной частью любой почки является конус нарастания, состоящий из меристематических клеток. Он даёт начало элементам структуры побега и корня, образует зачатки листьев, почек и др.
По наличию в них органов плодоношения различают бесплодные и плодоносные почки. Бесплодная почка несёт на зачаточном стебле эмбриональные листья, прилистники, усики и, развиваясь, образует бесплодный побег. Плодоносная почка несёт на зачаточном стебле, кроме указанных органов, эмбриональные соцветия и даёт начало плодоносному побегу.
По местоположению различают верхушечные (терминальные) почки, продолжающие рост главного побега, и боковые, обеспечивающие образование боковых побегов. Верхушечкая почка виноградного побега после вегетации отмирает, и на следующий год рост возобновляется за счёт боковых почек.
У винограда встречаются почки нескольких типов: почечка зародыша, пазушная пасынковая почка, центральная и замещающая почки зимующего глазка, зимующая пазушная почка, спящая почка.
При прорастании семени в процессе развития почечки зародыша (точка роста эпикотиля) образуется верхушечная почка сеянца. Развитие этой почки приводит к формированию первого побега стебля. В пазухе развивающихся на нём листьев закладываются пазушные почки: пасынковая и почки зимующего глазка. Верхушечная почка сеянца функционирует только в первый год, после перезимовки рост возобновляется за счёт почек зимующего глазка. Из почек зимующего глазка формируется также стебель саженца и ежегодно возобновляется рост растений винограда.
Пазушные почки винограда формируются в следующем порядке (рис. 15).
На растущей верхушке основного побега образуются бугорки листьев, которые по мере удаления от верхушки вытягиваются, в их пазухе закладывается пазушная пасынковая почка. Эта почка не имеет периода покоя, прорастает сразу же после закладки и даёт начало пасынку (рис. 16). В начале развития пасынкового побега в пазухе его первого листового зачатка закладывается новая почка, которая обычно не прорастает в текущую вегетацию, а только дифференцируется, превращаясь в сложную почку, так называемый зимующий глазок. Первый лист пасынка, как правило, остаётся в виде чешуйки.
Полностью сформировавшийся к концу вегетации глазок состоит из нескольких почек (рис. 17). В центре глазка располагается наиболее развитая центральная (главная) почка, а вокруг неё 3…6 замещающих (запасных). Эмбриональный побег главной почки имеет 9…14 узлов, на нижних из которых располагаются зачаточные бугорки будущих почек — зимующие пазушные почки. Замещающие почки менее дифференцированы, имеют 3…5 зачаточных узлов и, как правило, не несут в себе зачатков соцветий. Почки глазка имеют общее основание и общие твёрдые кутинизированные покровные чешуи. Почки плотно укутаны волосяным покровом. Почки зимующего глазка располагаются на узлах основного побега на возвышении, называемом подушечкой, которая соединена с диафрагмой. Между основанием почек и подушечкой находится тонкий слой паренхимных клеток, богатых хлорофиллом, — подстилающий слой. Существует мнение, что клетки подстилающего слоя способны формировать новые почки при повреждении глазка в период его формирования. Глазки, имеющие хотя бы в одной почке зачатки соцветий, называются плодоносными, не имеющие — бесплодными. Между плодоносными и бесплодными зимующими глазками отсутствуют внешние различия, для оценки эмбриональной плодоносности глазков проводятся в период покоя специальные исследования. Такие исследования необходимы для определения потенциальной плодоносности, прогнозирования и планирования урожая, обрезки.
Зимующие глазки формируются последовательно от основания к верхушке побега по мере его роста в течение вегетации. Для развития глазков, расположенных на разных узлах однолетнего побега, складываются различные внешние и внутренние условия: обеспеченность теплом, влагой, уровень облиственности, снабжение продуктами ассимиляции, интенсивность процессов метаболизма. Поэтому глазки разных узлов по длине однолетнего побега различаются по плодоносности. Как правило, в нижней части побега располагаются наименее плодоносные глазки, затем плодоносность их нарастает, достигает максимума и затем снижается. У самого основания однолетнего побега образуются 2…3 слабодифференцированных глазка, расположенных на сближенных междоузлиях, так называемые угловые глазки. Почки угловых глазков обычно бесплодные, но некоторые сорта обладают способностью формировать много плодоносных угловых глазков (Алиготе, Траминер, Московский устойчивый). Угловые глазки имеют практическое значение при слабом вызревании побегов, в условиях северного виноградарства. Побеги образующиеся из почек угловых глазков, используются для восстановления частей куста и формирования ассимилирующей поверхности при повреждении заморозками, градом. В зависимости от условий конкретного вегетационного года зона размещения наиболее продуктивных глазков смещается по длине однолетнего побега.
Эмбриональные побеги зимующего глазка из зачатка почки, заложившейся в пазухе первого листа пасынкового побега, формируются следующим образом. Сразу после своего появления эта почка имеет вид меритематического бугорка. В результате деления клеток в конусе нарастания увеличиваются её размеры. По мере удлинения главной оси меристематического бугорка сбоку от конуса нарастания последовательно отчленяются зачатки листьев (моноподиальное ветвление). По мере удаления от верхушки почки листовые зачатки вытягиваются, дифференцируются , а в пазухе нижних листьев закладываются зачатки почек следующей генерации. Рост эмбрионального побега центральной почки по моноподиальному типу происходит до 3…5-го узла, а затем он переходит к симподиальному (следующие два узла). На 3…5-м узле главная ось эмбрионального побега отклоняется в сторону и формирует зачаточное соцветие или усик, а новая точка роста, сформировавшаяся в пазухе верхнего листа, продолжает её рост (рис. 18). На следующем зачаточном узле также происходит рост по симподиальному типу — сменяется точка роста и образуется соцветие или усик. Затем образуется один зачаточный узел с моноподиальным типом ветвления. Таким образом, рост эмбриональных побегов почек зимующих глазков в процессе их дифференциации в год заложения происходит по характерному для виноградного растения симподиально-моноподиальному типу.
В морфогенезе почек зимующего глазка можно установить периоды, которые являются определяющими в формировании его плодоносности. Первый критический период — образование первого узла симподиального типа на зачаточном стебле центральной почки. Уровень обеспеченности и направленность процессов метаболизма в этот период, гормональный баланс определяют образование из отклонившейся главной оси эмбрионального побега зачаточного соцветия или усика. Если при этом образуется соцветие, то в следующую вегетацию из почки обычно развивается плодоносный побег. А если на первом узле симподиального типа эмбрионального побега формируется зачаточный усик, то в следующую вегетацию из него развивается бесплодный побег, так как выше усика соцветие образоваться у виноградного побега не может. Поэтому период формирования первого узла симподиального типа профессор А. М. Нергуль назвал «первым и главным критическим периодом в формировании генеративных органов». Условия формирования следующих нижних узлов симподиального типа при образовании на первом из них зачаточного соцветия будет определять, сколько соцветий образуется на побеге. Главный критический период для разных почек зимующего глазка, почек глазков разных узлов, зимующих почек разных побегов наступает в разное время. Для почек каждого растущего побега этот период наступает последовательно, начиная с нижних узлов, перемещаясь к верхним. Установлено также, что в центральных почках нижних узлов растущего побега (часть из них остаётся после обрезки перед следующей вегетацией) он наступает в период, близкий к фазе цветения соцветий текущего года.
После закладки бугорков соцветий в почках зимующего глазка до осени идёт их дифференциация. Главная ось зачаточного соцветия, которая формируется из отклонившейся верхушки эмбрионального побега, развивается по моноподиальному типу. По мере роста главной оси сбоку формируются зачатки чешуек, в пазухе которых закладываются меристематические бугорки, формирующие оси соцветия следующего порядка. Для последних также характерен моноподиальный тип ветвления, т. е. по мере роста их главной оси сбоку закладываются чешуйки, а в их пазухе — меристематические бугорки, дающие начало осям следующего порядка, и т. д. Как правило, ветвление зачаточного соцветия происходит до 3…4-го порядка и органичивается в год закладки почек образованием примордиев цветков. В период покоя дифференциация структуры соцветия происходит весной по мере распускания почек и роста побега его несущего — формируются элементы цветка и др.
Зачаток соцветия, расположенный в почке зимующего глазка, к осени имеет вид конусовидного образования. Степень дифференциации и размеры зачаточного соцветия зависят от сорта, природных условий, ухода. Если на эмбриональном побеге развивается несколько зачаточных соцветий, то наиболее развитое из них первое, затем степень дифференциации и размеры их снижаются по мере удаления от основания зачаточного побега, его несущего, прерывается при наступлении фазы глубокого покоя почек — в районах культуры винограда Северного полушария в августе.
Внешние и внутренние условия периода формирования на побегах текущего года почек нижних узлов определяются во многом количеством соцветий, которые развиваются на кустах винограда в следующем году.Наиболее важными из них являются сорт, погодные условия, уровень питания, влагообеспеченность, гормональное состояние растения и некоторые другие.
Интенсивная дифференциация элементов почек, плодоносность почек по длине побега, степень дифференциации соцветий в значительной степени определяются сортом. Установлено, что у представителей восточной группы сортов вида V. vinifera L. в одних и тех же условиях закладка соцветий в центральных почках зимующих глазков происходит с отставанием по сравнению с сортами западно-европейской эколого-географической группы и группы бассейна Чёрного моря. Наиболее плодоносные глазки у сортов восточной группы формируются выше от основания побега, поэтому они требуют более длинной обрезки.
Путём применения различных агротехнических приёмов можно направленно влиять на процесс закладки и дифференциации почек и зачаточных соцветий. Установлено положительное влияние температуры на этот процесс. Хорошая освещённость внутри кроны куста стимулирует процесс фотосинтеза, улучшает питание развивающихся почек и повышает плодоносность их. Наряду с этим установлено положительное влияние солнечной радиации на морфогенез глазка. Затемнение глазков в период их формирования ухудшает закладку соцветий и степень их дифференциации (А. П. Дикань).
На плодоносность почек влияет интенсивность роста побегов. У слаборастущих побегов закладка соцветий и их дифференциация идёт слабее, чем у сильнорастущих. Поэтому на плодоношение при обрезке оставляют сильные побеги. Вместе с тем побеги, обладающие чрезмерно сильным ростом — жирующие побеги, характеризуются низкой плодоносностью, наиболее плодоносные глазки у них располагаются выше от основания побега. Более высокая плодоносность почек, оптимальное размещение плодовых почек по длине однолетнего побега наблюдается у нормальных, полноценных побегов. Свободное свисание побегов способствует повышению плодоносности почек, расположенных у основания. Установлено также, что кратковременное прекращение роста побегов путём прищипывания перед цветением стимулирует закладку соцветий в нижней зоне побега. Обработка кустов винограда перед цветением растворами ретардантов тормозит рост побегов и стимулирует закладку соцветий в почках нижних узлов.
Установлено также благоприятное влияние на формирование плодоносных почек минерального питания, микроэлементов (бора, марганца, цинка, молибдена). Обнаружена положительная связь между концентрацией крахмала в тканях побегов и закладкой зачаточных соцветий в зимующих глазках. Рядом исследователей определено, что существенно влияют не процесс закладки соцветий операции с зелёными частями куста.
Пасынковая пазушная почка прорастает в пазухах листьев каждого узла основных побегов сразу после своей закладки. При нормальной и повышенной нагрузке основными побегами пасынки развиваются слабо, чаще, чуть развившись, отсыхают и отпадают. При сильном росте пасынка на его узлах в пазухе листьев закладываются пазушные почки, из которых развиваются пасынки следующего порядка. При хорошем росте этих пасынков у основания их в пазухе первых листьев — чешуй закладываются новые почки, которые развиваются в зимующие глазки, расположенные на узле пасынка. Зимующие глазки, образовавшиеся на узлах пасынка, имеют такое же строение, что и зимующие глазки основных побегов. Также они могут быть плодоносными и бесплодными. При сильном нарушении корреляции между корневой системой и надземной частью могут развиваться пасынки и зимующие глазки следующих порядков (рис. 19).
Почки зимующего глазка развиваются в обычных условиях только после перезимовки. Обычно развивается центральная почка и образует побег. Если повреждается центральная почка, то развивается наиболее дифференцированная замещающая, при её повреждении — следующая по степени дифференциации. Когда развиваются центральная и одна из замещающих почек, образуются побеги-двойники. Иногда развиваются три почки зимующего глазка, формируя побеги-тройники. Это бывает при нарушении корреляции между подземной и надземной частями растения из-за повреждения части куста или части почек. Иногда не развивается ни одна из почек глазка. Не проросшие на следующий после своего образования год живые центральная и замещающая почки выпадают, а на их месте остаётся почечных след — «спящая почка», которая сохраняется на стебле два и более года. По мере утолщения стебля происходит и заглубление в древесину спящей почки. Эти почки сохраняют жизнеспособность в течение десятков лет. Они развиваются при нарушении корреляции между надземной и подземной частями куста и образуют порослевые и волчковые побеги. Роль спящих почек велика для омоложения и восстановления отдельных частей куста: рожков, рукавов, штамба, иногда всей надземной части куста. Порослевые и волчковые побеги используют также для укладки отводков.
Таким образом, виноградное растение в результате эволюции выработало способность к образованию большого разнообразия почек — пазушных (пасынковых), главных, замещающих, спящих (рис. 19). Существует определённая, контролируемая самим растением последовательность в развитии почек. В первую очередь развиваются пазушные пасынковые, затем центральная, замещающая, а в последнюю очередь — спящая. Раньше развивается более дифференцированная почка, затем следующая по степени дифференциации и т. д. По мнению профессора А. М. Негруля, это связано с тем, что разным почкам нужны различного уровня питание и корневое давление. Большую роль в развитии почек, выхода их из состояния относительного покоя играют фитогормоны.
Всё это обеспечивает высокую степень устойчивости виноградного растения к неблагоприятным условиям среды, возможность восстановления после повреждения.
Схематическое изображение циклов развития пазушных почек представлено на рис. 19.